Estudo apresenta descobertas valiosas sobre o envolvimento do sistema endocanabinoide na progressão da endometriose
Descrição
Endocanabinoides e seus receptores modulam a patogênese da endometriose e a resposta imune (https://doi.org/10.1101/2023.10.11.561929) A endometriose (EM), caracterizada pela presença de tecido semelhante ao endométrio fora do útero, é a principal causa de dor pélvica crônica e infertilidade em mulheres em idade reprodutiva. Apesar de sua alta prevalência, os mecanismos moleculares subjacentes à patogênese do EM permanecem pouco compreendidos. O sistema endocanabinoide (ECS) é conhecido por influenciar várias características cardinais dessa doença complexa, incluindo dor, vascularização e sobrevida global da lesão, mas os mecanismos exatos não são conhecidos. Utilizando modelos murinos de EM knockout para CNR1 (k/o), CNR2 k/o e tipo selvagem (WT), revelamos as contribuições do ECS e desses receptores na iniciação da doença, progressão e modulação imune. Particularmente, identificamos disfunção de células T específicas para EM no modelo murino de EM CNR2 k/o. Também demonstramos o impacto das alterações induzidas pela decidualização nos componentes do SEC e o panorama transcricional único associado à doença dos componentes do SEC no EM. A análise por Citometria de Massa (IMC) por imagem revelou características distintas do microambiente entre os genótipos CNR1, CNR2 e WT na presença ou ausência de decidualização. Este estudo, pela primeira vez, fornece uma análise aprofundada do envolvimento do SEC na patogênese do EM e estabelece as bases para o desenvolvimento de novas intervenções terapêuticas para aliviar o impacto desta condição debilitante.
Avaliação eLife
Este estudo apresenta descobertas valiosas sobre o envolvimento do sistema endocanabinoide na progressão da endometriose, utilizando modelos murinos knockout (KO) para CNR1 e CNR2. As evidências que sustentam as alegações dos autores são incompletas ; a inclusão de sequenciamento de RNA em massa, citometria de fluxo e citometria de massa por imagem teriam fortalecido o estudo. Este trabalho pode ser de interesse para cientistas médicos que trabalham com endometriose.
https://doi.org/10.7554/eLife.96523.3.sa0
https://elifesciences.org/articles/96523
https://elifesciences.org/articles/96523
Resumo
A endometriose (EM), caracterizada pela presença de tecido semelhante ao endométrio fora do útero, é a principal causa de dor pélvica crônica e infertilidade em mulheres em idade reprodutiva. Apesar de sua alta prevalência, os mecanismos moleculares subjacentes à patogênese da EM permanecem pouco compreendidos. Sabe-se que o sistema endocanabinoide (SE) influencia diversas características cardinais dessa doença complexa, incluindo dor, vascularização e sobrevida global da lesão, mas os mecanismos exatos não são conhecidos. Utilizando modelos murinos de EM com knockout para CNR1 (k/o), CNR2 k/o e tipo selvagem (WT), revelamos as contribuições do SE e desses receptores na iniciação, progressão e modulação imunológica da doença. Em particular, identificamos disfunção de células T específicas para SE no modelo murino de EM com CNR2 k/o. Também demonstramos o impacto das alterações induzidas pela decidualização nos componentes do SE e o panorama transcricional único associado à doença dos componentes do SE no EM. A análise por citometria de massa (IMC) por imagem revelou características distintas do microambiente entre os genótipos CNR1, CNR2 e WT na presença ou ausência de decidualização. Este estudo, pela primeira vez, fornece uma análise aprofundada do envolvimento do SEC na patogênese do EM e estabelece as bases para o desenvolvimento de novas intervenções terapêuticas para aliviar o impacto desta condição debilitante.
Introdução
O EM é um distúrbio ginecológico crônico caracterizado pela presença e crescimento de tecido semelhante ao endométrio fora do útero, denominado lesões ectópicas. Apesar de seu impacto global em aproximadamente 200 milhões de indivíduos e da profunda redução em sua qualidade de vida, as origens exatas do EM permanecem indefinidas ( Giudice e Kao, 2004 ). Evidências acumuladas, incluindo nossos estudos anteriores, destacam que componentes do SEC são desregulados dentro do microambiente da lesão do EM, bem como na circulação sistêmica de pacientes com EM ( Lingegowda et al., 2021b ; Bilgic et al., 2017 ; Shen et al., 2019 ). O SEC é uma rede de sinalização complexa composta por receptores canônicos (CNR1 e CNR2) e ligantes endocanabinoides (EC), juntamente com uma rede de sinalização estendida não canônica de ligantes e enzimas extensivamente revisada em outro lugar ( Di Marzo et al., 2004 ). CNR1 e CNR2 são expressos principalmente em tecidos nervosos, células imunes e tecidos reprodutivos, onde regulam vários processos fisiológicos, incluindo percepção da dor, respostas imunes e funções reprodutivas ( Zou e Kumar, 2018 ). Consequentemente, a patogênese da EM foi postulada como consequência da deficiência de EC ( Russo, 2016 ; Lingegowda et al., 2022 ).
Embora a etiologia precisa do EM não seja conhecida, a teoria amplamente aceita de Sampson sobre menstruação retrógrada sugere que as lesões de EM se originam de fragmentos endometriais refluídos depositados durante a menstruação (Sampson, 1927) . Tanto a gravidez quanto a menstruação dependem da decidualização espontânea do estroma endometrial, que é extensivamente remodelado sob a influência de hormônios, fatores de crescimento e citocinas selecionadas que orquestram o recrutamento de células imunes e adaptações vasculares ( Gellersen e Brosens, 2014 ). Há evidências claras de que pacientes com EM apresentam defeitos no endométrio eutópico, incluindo expressão diferencial de marcadores-chave de receptividade endometrial, como fator inibitório de leucemia ( LIF ), proteína arginina metiltransferase 5 ( PRMT5 ) e proteína homeobox hox-A10 ( HOXA10 ), que foram associados ao EM e subsequente infertilidade ( Cai et al., 2022 ; Tomassetti e D'Hooghe, 2018 ; Zutautas et al., 2023 ). As evidências também sugerem que componentes do ECS, incluindo CNR1 e CNR2, são importantes para manter a integridade do tecido durante a decidualização e a implantação bem-sucedida do embrião. De fato, vários relatórios indicam que camundongos sem receptores canabinoides, CNR1 e CNR2, apresentaram implantação prejudicada, aumento nas taxas de falha na gravidez, edema elevado e formação inadequada da zona decidual primária, destacando o papel crucial da sinalização do ECS na decidualização, implantação e gravidez bem-sucedidas ( Wang et al., 2008 ; Li et al., 2019 ; Sun et al., 2010 ).
No ME, a ativação de CNR1 e CNR2 auxilia no controle da proliferação de lesões, dor e vascularização ( Lingegowda et al., 2021a ; Leconte et al., 2010 ). Considerando a sinalização desregulada do SEC e seu papel central na decidualização e fertilidade, levantamos a hipótese de que a expressão alterada de CNR1 e CNR2 interromperá a dinâmica de sinalização do SEC, levando ao desenvolvimento de lesões. Além disso, o envolvimento do SEC na modulação da resposta imune e da homeostase pode interromper a dinâmica imune e promover o estabelecimento de lesões.
Conduzimos uma investigação abrangente sobre o papel do SEC desregulado no estabelecimento e progressão do EM utilizando modelos murinos CNR1 k/o e CNR2 k/o. Para abordar as causas subjacentes da disfunção do SEC, induzimos a decidualização artificial em camundongos WT, CNR1 k/o e CNR2 k/o e usamos fragmentos endometriais de cornos uterinos decidualizados (DD) e undecidualizados (UnD) para induzir o EM em camundongos receptores de seus respectivos genótipos. Além disso, exploramos o potencial imunomodulador do SEC no EM, esclarecendo como alterações na sinalização do EC podem influenciar o comportamento das células imunes dentro do meio peritoneal localizado em camundongos induzidos com EM. Nosso estudo contribui para o conhecimento fundamental em torno da desregulação do SEC no EM e abre caminho para potenciais estratégias terapêuticas direcionadas ao SEC para o manejo da doença.
Resultados
Ligantes do ECS são desregulados em um modelo murino de EM sem receptores CNR1 e CNR2
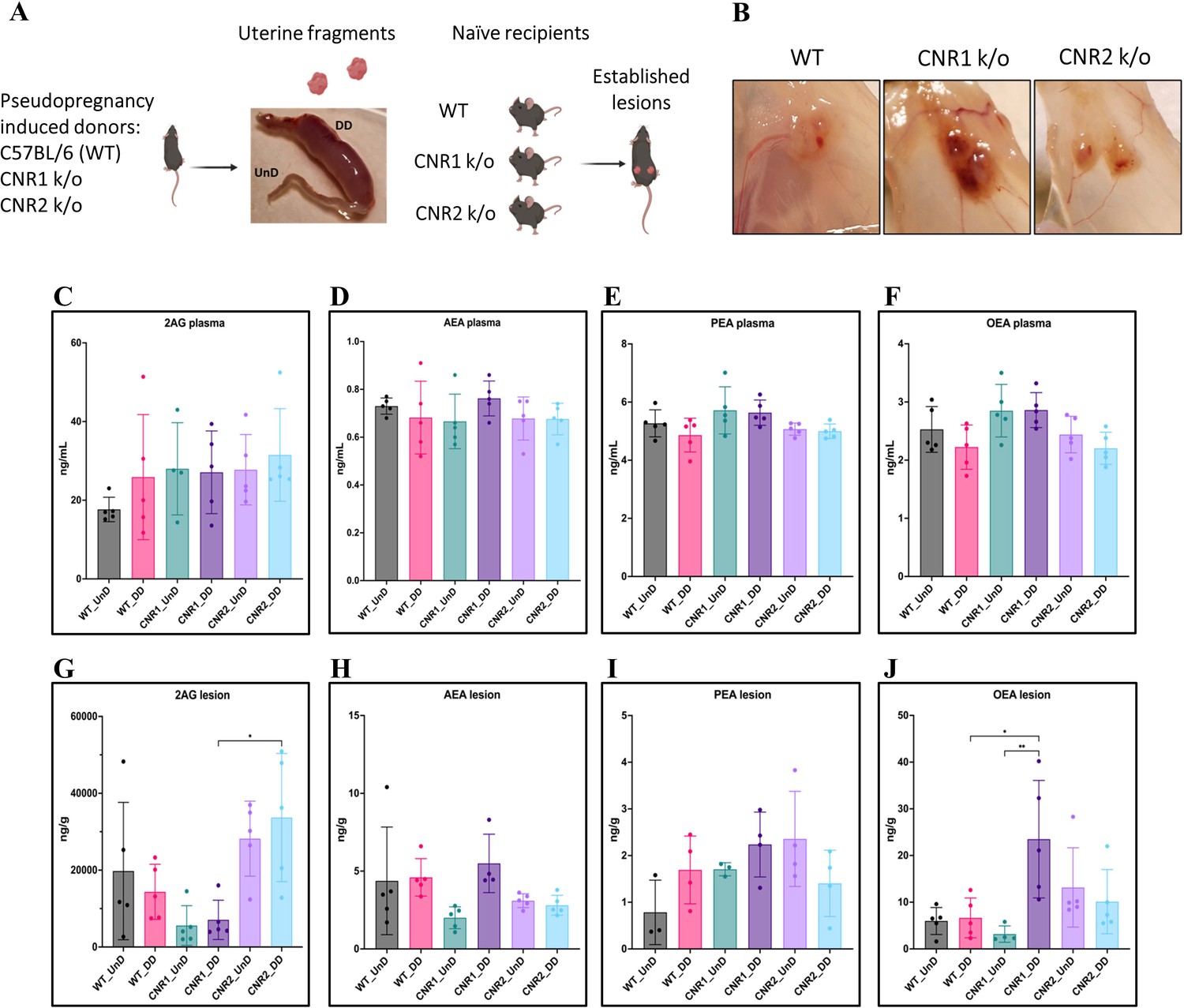
Com base em nosso trabalho anterior demonstrando ligantes desregulados do ECS em pacientes e em nosso modelo murino de EM ( Lingegowda et al., 2021b ), primeiro avaliamos se a ausência de CNR1 ou CNR2 levou a alterações no ligante do ECS. Para isso, realizamos espectrometria de massa direcionada em lesões plasmáticas e de EM obtidas de CNR1 k/o, CNR2 k/o e seus controles WT. Nesses camundongos, o EM foi induzido usando tecidos UnD e DD obtidos de suas respectivas linhagens em receptores compatíveis (por exemplo, UnD e DD de camundongos doadores CNR1 k/o foram implantados em camundongos receptores CNR1 k/o). Detectamos alguns dos principais ligantes de EC, como 2-araquidonoilglicerol (2-AG), N-araquidonoiletanolamina (AEA), palmitoiletanolamida (PEA) e oleoiletanolamida (OEA) em lesões plasmáticas e de EM de todos os genótipos. Todos os ligantes ECS identificados são predominantemente anti-inflamatórios e a gama de 2-AG, AEA, PEA e OEA no plasma e lesões foram comparáveis ao nosso estudo anterior ( Lingegowda et al., 2021b ). No plasma, não encontramos diferenças significativas nos ligantes ECS em todos os grupos ( Figura 1C–F ), o que pode ser devido à rápida homeostase alcançada na circulação ( Lu e Mackie, 2016 ). No entanto, no microambiente da lesão, capturamos níveis mais altos de vários ligantes EC ( Figura 1G–J ). Em camundongos CNR1 k/o, concentrações significativamente maiores de OEA foram observadas nas lesões DD em comparação com as lesões UnD ( Figura 1J ) e, no geral, foi em média duas vezes maior em comparação com ambas as lesões de camundongos WT e CNR2 k/o. O 2-AG, que se liga seletivamente ao receptor CNR2, apresentou valores significativamente mais elevados nas lesões de EM de UnD e DD em camundongos CNR2 k/o ( Figura 1G ), em comparação com os camundongos CNR1 k/o. Isso pode indicar uma resposta compensatória na ausência de CNR2. Em conjunto, esses achados fornecem insights sobre a potencial desregulação dos ligantes do ECS na ausência de CNR1 e CNR2 e seu envolvimento no cenário DD vs. UnD durante o estabelecimento de lesões de EM.
Caracterização de ligantes endocanabinoides em um modelo murino singênico modificado de endometriose.
( A ) Visão geral do modelo murino singênico modificado de endometriose (EM) onde camundongos pseudográvidos selvagens (WT), CNR1 k/o e CNR2 k/o foram induzidos com decidualização (DD) pela injeção de óleo de gergelim…
Impacto na expressão gênica e alterações na via em lesões de EM de camundongos na ausência de CNR1 e CNR2
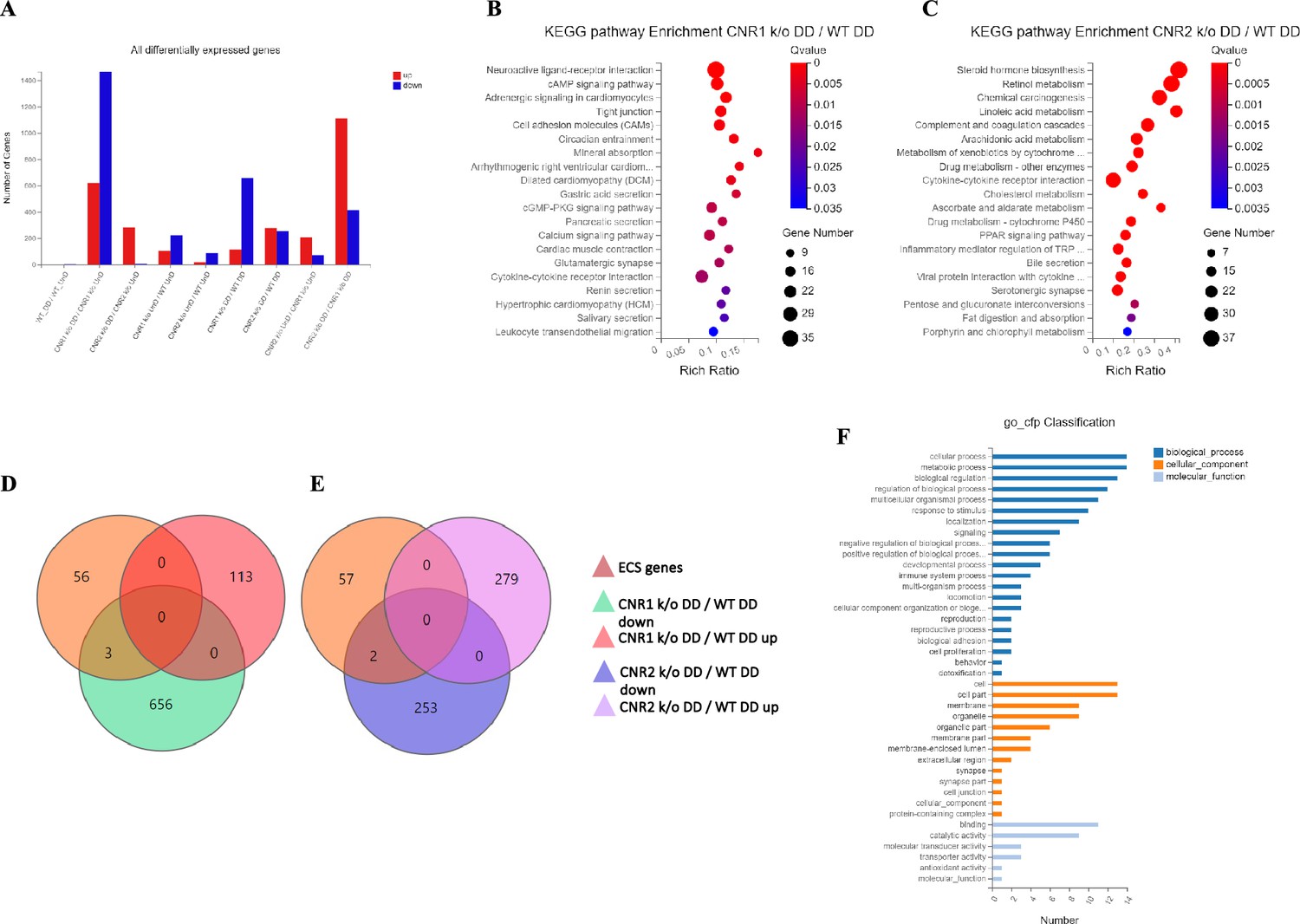
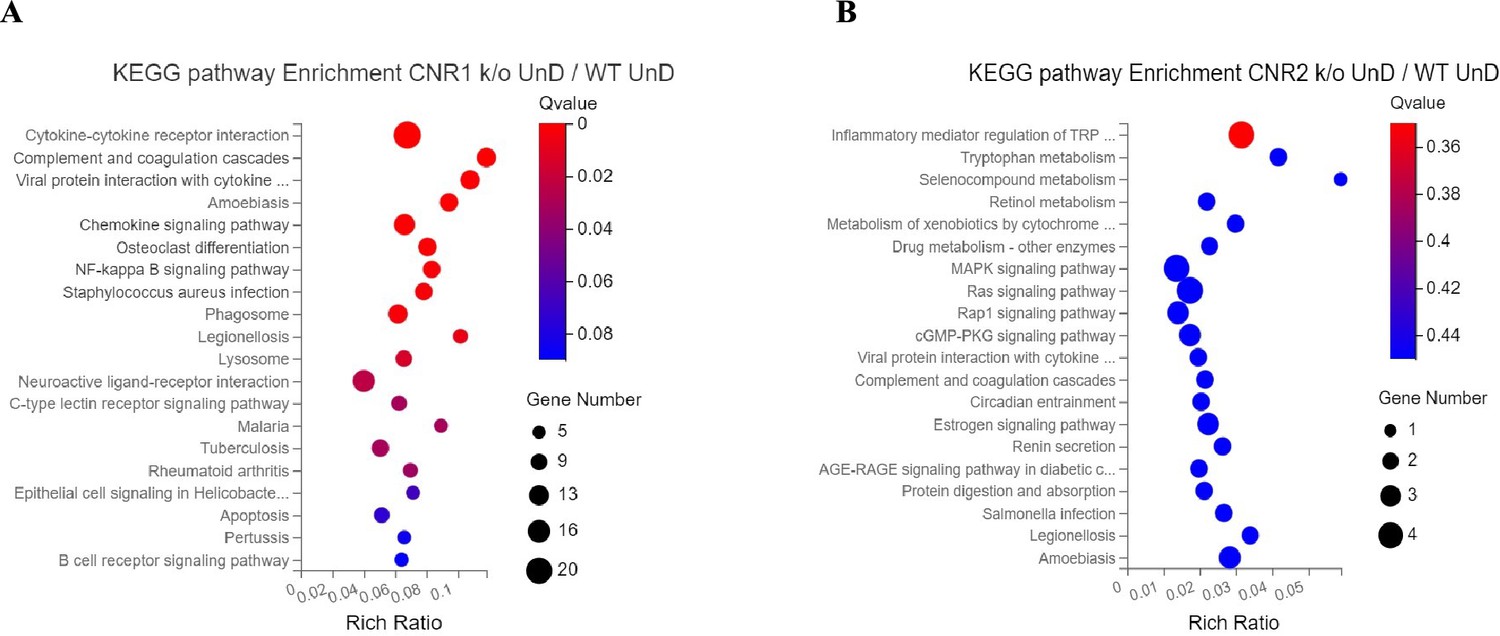
Em seguida, investigamos os efeitos da ausência de CNR1 e CNR2 no perfil transcriptômico de lesões EM de UnD e DD de seus respectivos genótipos. O sequenciamento de RNA em massa foi realizado em lesões de UnD e DD de camundongos WT, CNR1 k/o e CNR2 k/o, conforme detalhado anteriormente, para elucidar as alterações moleculares associadas à interrupção da sinalização desses dois receptores. A análise de expressão diferencial revelou alterações nos perfis de expressão gênica entre os diferentes genótipos e tipos de lesão ( Figura 2A ). Um total de 1100 e 639 genes diferencialmente expressos (DEGs) foram encontrados em lesões de UnD e DD de camundongos CNR1 k/o e CNR2 k/o, respectivamente, em comparação aos controles WT (os dados de UnD são fornecidos no arquivo suplementar 2 ). Para obter insights sobre as implicações biológicas das mudanças observadas na expressão gênica, conduzimos uma análise de enriquecimento da via da Enciclopédia de Genes e Genomas de Kyoto (KEGG) nos DEGs identificados em lesões UnD e DD de camundongos CNR1 k/o e CNR2 k/o em comparação com camundongos WT. Nas lesões DD de camundongos CNR1 k/o, a análise da via KEGG revelou alterações significativas em várias vias ( Figura 2B ). Notavelmente, a via das moléculas de adesão celular foi proeminentemente afetada, indicando um papel potencial para CNR1 na mediação de interações célula-célula e processos de remodelação de tecidos. Além disso, a sinalização do monofosfato de adenosina cíclico (AMPc) emergiu como outra via impactada negativamente, implicando CNR1 na modulação de cascatas de sinalização intracelular. Em lesões DD de camundongos CNR2 k/o, a análise destacou vias distintas afetadas no contexto de inflamação e EM ( Figura 2C ), incluindo a via de interações do receptor de citocina, apontando para o envolvimento de CNR2 em respostas imunes e processos inflamatórios associados ao EM. Além disso, capturamos alterações na via de biossíntese do hormônio esteroide, sugerindo um papel para o CNR2 em mecanismos relacionados a hormônios relevantes para o desenvolvimento e homeostase do tecido endometrial.
O perfil transcriptômico de lesões semelhantes à endometriose em camundongos com receptores canônicos desativados (CNR1 e CNR2) revela extensa expressão genética diferencial e vias alteradas.
( A ) Resumo dos genes diferencialmente expressos (DEGs) da análise de sequenciamento de RNA em massa conduzida em lesões não decidualizadas (UnD) e decidualizadas (DD) de tipo selvagem (WT), CNR1 k/o e …
Genes diferencialmente expressos (DEGs) de sequenciamento de RNA em massa entre as lesões não decidualizadas (UnD) de CNR1 k/o e CNR2 k/o em comparação com controles do tipo selvagem (WT).
( A, B ) Os gráficos de vulcão para lesões decidualizadas (DD) de CNR1 k/o e CNR2 k/o em comparação com WT, respectivamente. A variação de log2 é representada no eixo x e o valor Q de log10 (ajustado por FDR) como...
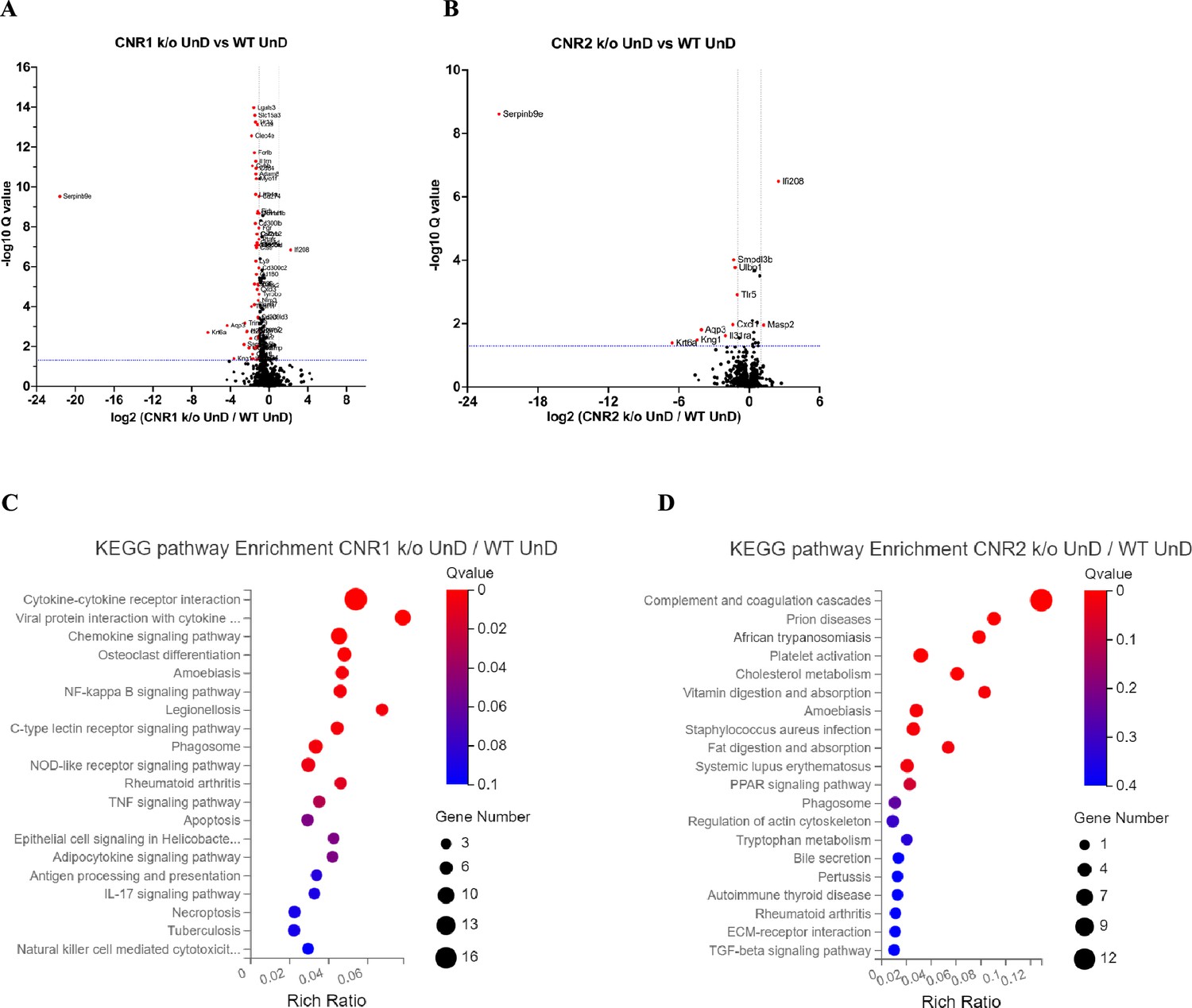
Em seguida, realizamos uma análise de subconjunto para genes diretamente envolvidos na sinalização do ECS. Um total de 59 genes-chave na sinalização do ECS foram selecionados, conforme identificado em um estudo de Tanaka et al., 2022. Surpreendentemente, apesar do papel central do CNR1 e do CNR2 na sinalização do ECS, encontramos um número limitado de DEGs relacionados a esse sistema. Dos 59 genes diretamente associados ao ECS, apenas 3 ( Cnr1, Plch1 e Plch2 ) e 2 ( Cnr2 e Plag2ge ) DEGs foram identificados nas lesões DD de CNR1 k/o ( Figura 2D ) e CNR2 k/o ( Figura 2E ), respectivamente, em comparação com as lesões DD de controles WT. Esse resultado sugere que CNR1 e CNR2 modulam o microambiente EM por meio de interações intrincadas com outras vias de sinalização além da via canônica do ECS. Uma análise abrangente de classificação de ontologia genética (GO) nos 59 genes ECS identificados ( Figura 2F ) revelou seus papéis multifacetados em funções reprodutivas, regulação do sistema imunológico e processos celulares. Os genes exibiram enriquecimento em funções moleculares, como atividade do receptor e ligação lipídica. Em termos de componentes celulares, esses genes foram associados a estruturas da membrana plasmática e compartimentos intracelulares, significando sua localização subcelular diversa e envolvimento potencial em processos celulares dinâmicos. Além disso, a análise GO destacou sua participação em processos biológicos, como modulação da resposta imune, metabolismo lipídico, comunicação celular e vias de sinalização intracelular, indicando o impacto mais amplo do ECS além de suas funções canônicas. Os resultados das comparações UnD entre os genótipos são fornecidos nos dados suplementares ( Figura 2—suplemento da figura 1 ). Embora as lesões UnD exibissem padrões distintos de expressão gênica em comparação com as lesões DD, tendências comuns nos efeitos de CNR1 k/o e CNR2 k/o na expressão gênica foram observadas em ambos os tipos de lesão. Especificamente, a interação do receptor de citocina, a cascata do complemento e as vias mediadoras inflamatórias foram significativamente alteradas nas lesões UnD de camundongos CNR1 k/o e CNR2 k/o, em comparação com as lesões UnD de camundongos WT. De modo geral, nossos achados destacam o impacto significativo de CNR1 e CNR2 k/o na expressão gênica em lesões de EM e as implicações para a patogênese do EM.
Interrupção de genes relacionados à resposta imune adaptativa em lesões de EM sem CNR1 e CNR2
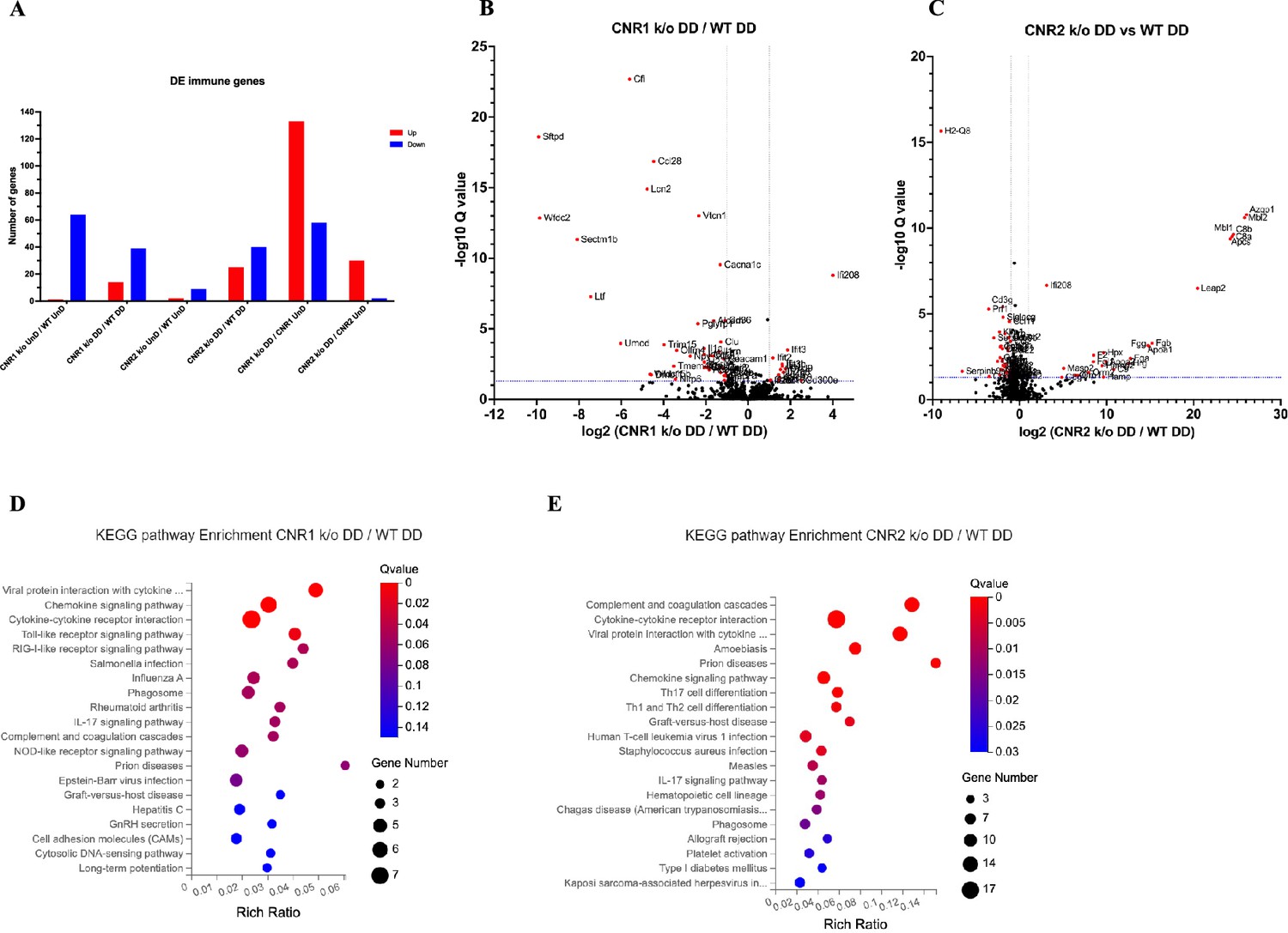
Com base em nossa investigação anterior sobre as alterações transcriptômicas, conduzimos uma análise aprofundada de genes relacionados ao sistema imunológico expressos diferencialmente (conforme InnateDB versão 5.4) em lesões UnD e DD em todos os genótipos. Aqui, nossa análise é focada nos genes relacionados ao sistema imunológico dentro de lesões DD de camundongos CNR1 k/o e CNR2 k/o em comparação com controles WT. A comparação das lesões UnD de CNR1 k/o e CNR2 k/o com camundongos EM WT está incluída nos dados suplementares ( Figura 3—suplemento da figura 1 ). O gráfico de barras de expressão diferencial ( Figura 3A ) fornece uma representação dos genes regulados positivamente e negativamente em cada comparação. Os gráficos de vulcão para lesões DD de CNR1 k/o vs. WT ( Figura 3B ) e CNR2 k/o vs. WT ( Figura 3C ) ilustram os vários DEGs. Notavelmente, as lesões CNR1 k/o DD exibiram 39 genes regulados negativamente (por exemplo, Nlrp6 e Il1a, reguladores essenciais da resposta inflamatória) e 14 genes regulados positivamente (por exemplo, Cxcl9 e Cxcl10, quimiocinas envolvidas no recrutamento de células imunes), enquanto as lesões CNR2 k/o DD mostraram 40 genes regulados negativamente (por exemplo, Siglecg e Il6, envolvidos na regulação imune e resposta pró-inflamatória) e 25 genes regulados positivamente (por exemplo, C8a , C9 e Masp2 , parte do sistema do complemento), destacando mudanças substanciais na expressão gênica associada à interrupção de CNR1 e CNR2 em comparação às lesões DD de camundongos WT. De particular interesse, observamos uma regulação negativa significativa de genes relacionados a células T ( Cd3e , Cd3g , Gata3 e Ctla4 ) nas lesões DD de CNR2 k/o ( arquivo suplementar 3 ), alinhando-se com a disfunção de células T CD3 + observada no PF e no baço e validações adicionais do ensaio funcional in vitro (mencionado abaixo). No entanto, não encontramos as mesmas diferenças (genes relacionados a células T) nas lesões UnD de camundongos CNR2 k/o. Além disso, as lesões UnD de camundongos CNR2 k/o mostraram um número significativamente baixo de DEGs (11 em comparação com 65 nas lesões DD de camundongos CNR2 k/o), sugerindo uma resposta dependente de decidualização ( arquivo suplementar 3). Esta observação enfatiza claramente uma ligação potencial entre a disfunção do CNR2 com a decidualização caracterizada por problemas de sinalização de células T dentro do microambiente EM. Para entender as implicações funcionais dos DEGs, conduzimos a análise da via KEGG em genes imunes específicos diferencialmente expressos em lesões DD de camundongos CNR1 k/o e CNR2 k/o. Notavelmente, em lesões DD CNR1 k/o comparadas com DD WT, a via de sinalização de quimiocina, a interação receptor de citocina-citocina e as vias de sinalização do receptor Toll-like (TLR) foram afetadas negativamente ( Figura 3D ). Essas descobertas fornecem insights sobre o impacto da interrupção do CNR1 na função das células imunes dentro do ambiente DD. Por outro lado, as lesões DD CNR2 k/o foram associadas a alterações significativas na interação receptor de citocina-citocina, vias de diferenciação de células Th17, Th1 e Th2 ( Figura 3E ). Essas vias comprovadamente estão envolvidas no desenvolvimento, diferenciação e funções efetoras das células T, em consonância com nossa desregulação observada de genes relacionados às células T. Em conjunto, essas descobertas elucidam ainda mais os papéis de CNR1 e CNR2 na modulação das respostas imunes no contexto do EM.
O sequenciamento de RNA em massa revelou alterações na expressão gênica relacionada ao sistema imunológico e na via em lesões de endometriose (EM) de camundongos CNR1 k/o e CNR2 k/o.
( A ) Visão geral do gráfico de barras dos genes relacionados ao sistema imunológico expressos diferencialmente (DE) entre diferentes genótipos e tipos de lesão. ( B, C ) Os gráficos de vulcão para lesões decidualizadas (DD) de CNR1 k/o vs. …
Análise da via da Enciclopédia de Genes e Genomas de Kyoto (KEGG) de genes diferencialmente expressos (DEGs) específicos do sistema imunológico.
( A, B ) Genes imunoespecíficos que foram expressos diferencialmente foram submetidos à análise da via KEGG entre lesões CNR1 k/o não decidualizadas (UnD) e lesões CNR2 k/o UnD em comparação com o tipo selvagem…
Citometria de fluxo multiespectral revelou perfis alterados de células imunes em um modelo murino de EM sem CNR1 e CNR2
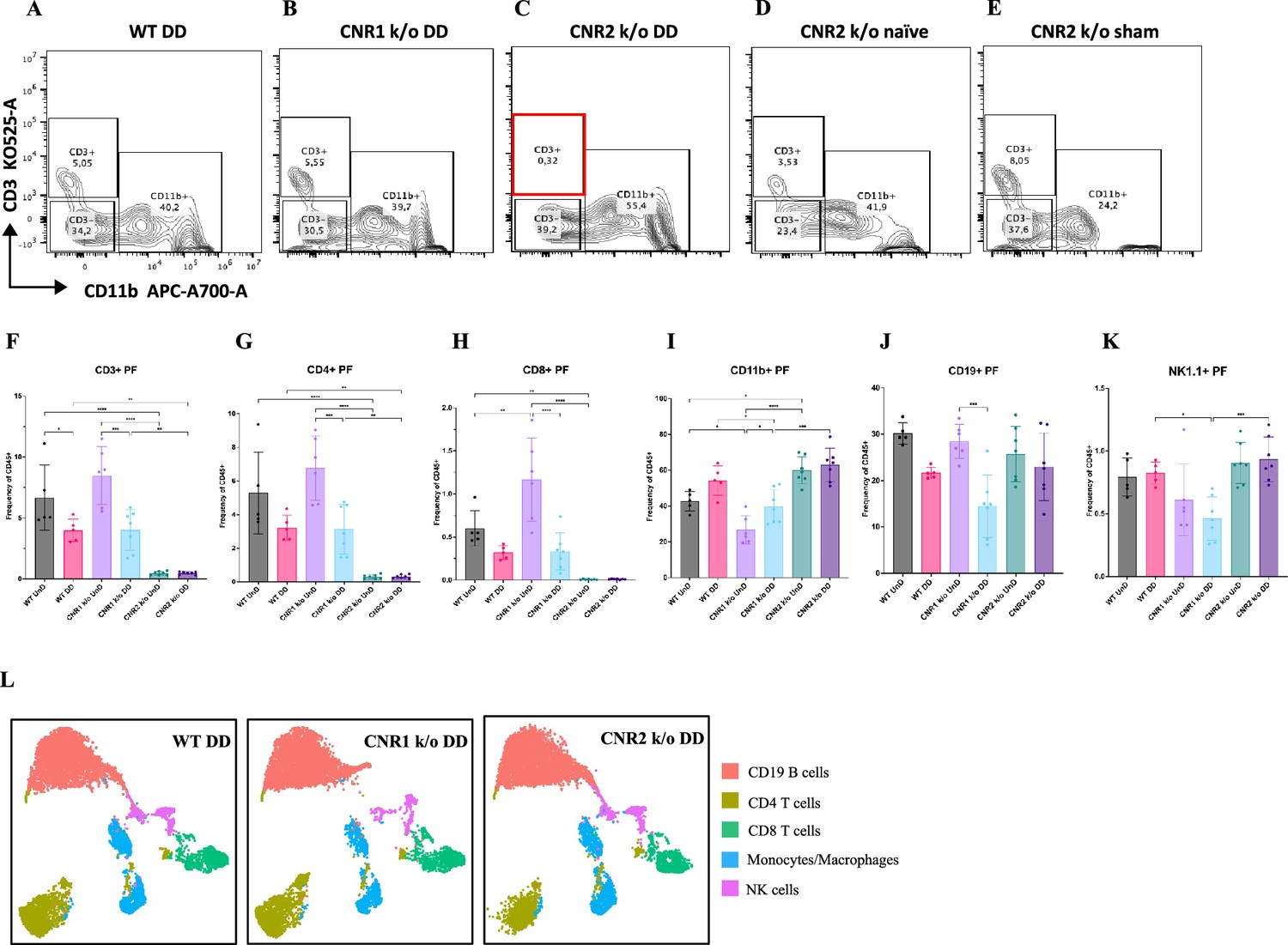
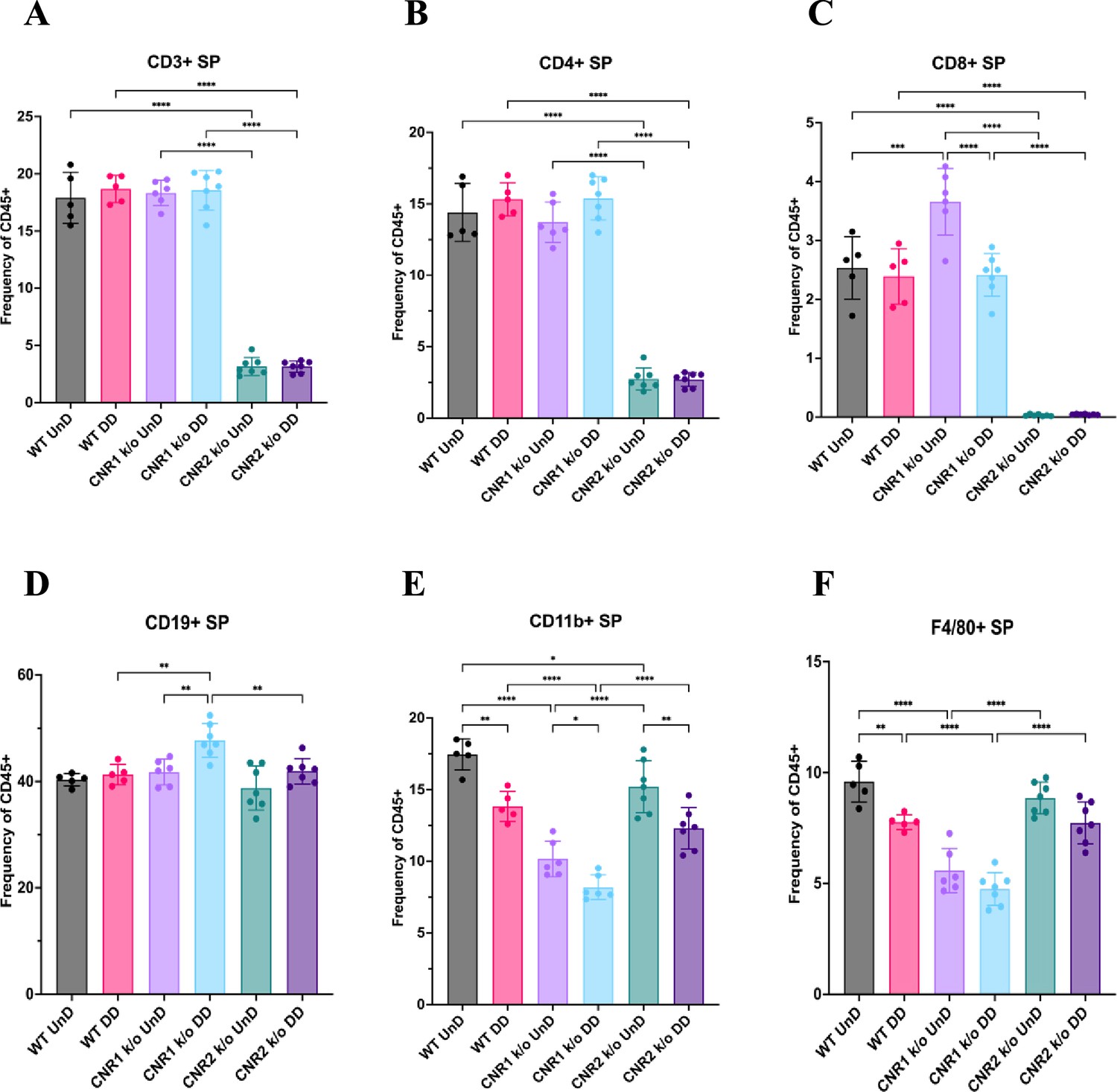
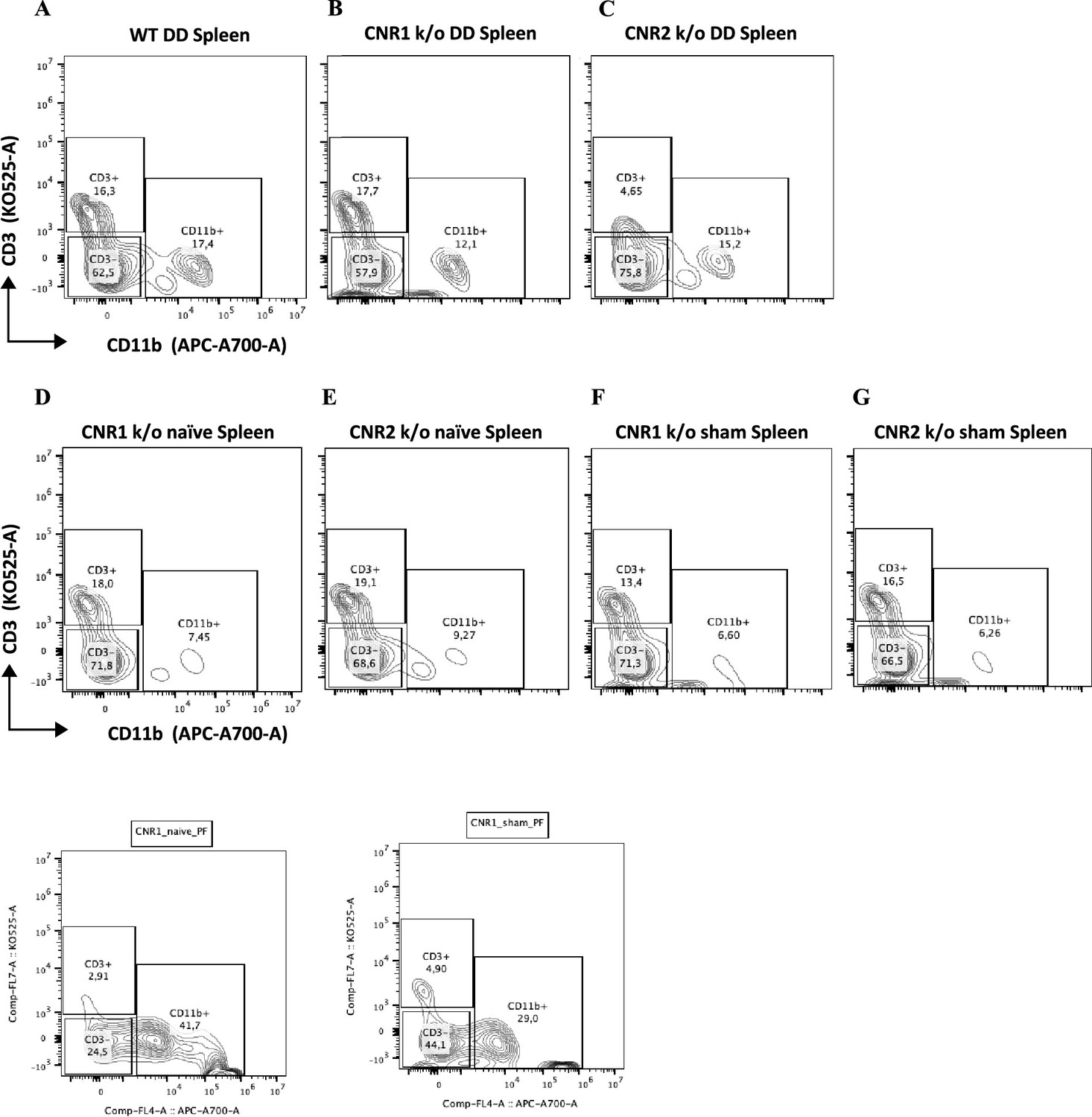
A desregulação imunológica é reconhecida como um fator crucial na patogênese do EM ( Symons et al., 2018 ). Para elucidar o impacto da ausência de CNR1 e CNR2 nas populações de células imunes, realizamos perfis imunes multiespectrais em esplenócitos e células do PF de camundongos WT, CNR1 k/o e CNR2 k/o portadores de lesões UnD e DD. Painéis de controle representativos de células CD3 (eixo y) vs CD11b (eixo x) do PF ( Figura 4A–E ) ilustram perfis distintos entre camundongos WT, CNR1 k/o e CNR2 k/o com EM, bem como camundongos CNR2 k/o naïves, não operados, e controles CNR2 k/o operados de forma simulada. Surpreendentemente, as populações de células T CD3 + estavam quase ausentes no PF de camundongos CNR2 k/o com EM, independentemente dos tipos de lesão (UnD e DD), quando comparados a outros grupos ( Figura 4C e F ). Essa tendência se estendeu ainda mais às células T auxiliares CD4 + e às células T citotóxicas CD8 + ( Figura 4G–H ). Camundongos CNR1 k/o com lesões DD exibiram frequências de células T CD3 + ( Figura 4F ), CD4 + ( Figura 4G ) e CD8 + ( Figura 4H ) significativamente reduzidas em comparação com suas contrapartes UnD, bem como populações menores de células B CD19 + e células NK1.1 + ( Figura 4K ), em comparação com camundongos WT e CNR2 k/o. Concomitantemente com a redução de subconjuntos de células T, um aumento em monócitos CD11b foi observado no PF de camundongos CNR2 k/o com lesões UnD e DD em comparação com camundongos WT e CNR1 k/o ( Figura 4I ). Similarmente, camundongos CNR1 k/o com lesões DD apresentaram populações maiores de monócitos/macrófagos em comparação com suas contrapartes UnD ( Figura 4I ). Além disso, camundongos WT com lesões DD demonstraram frequências de células T CD3 + significativamente menores em comparação com suas contrapartes UnD ( Figura 4F ), sugerindo um efeito associado à decidualização. Os esplenócitos exibiram tendências análogas ( Figura 4L ), conforme representado pelos gráficos tSNE (gráficos de barras e gráficos de contorno na Figura 4 - suplementos de figura 1 e 2 ). Essas alterações no número de células imunes reforçam a influência da desregulação e decidualização de CNR1 e CNR2 nas populações de células imunes, confirmadas tanto localmente no PF quanto sistemicamente no baço.
O perfil de citometria de fluxo do fluido peritoneal (FP) e das células esplênicas mostra fenótipos alterados de células imunes em camundongos CNR1 k/o e CNR2 k/o com endometriose (EM).
( A–E ) Painéis de gating de células PF mostrando CD3 KO525-A no eixo y vs CD11b APC-A700-A no eixo x entre camundongos selvagens (WT), CNR1 k/o e CNR2 k/o com lesões EM decidualizadas (DD), bem como…
Análise de citometria de fluxo de fenótipos de células imunes em esplenócitos de camundongos selvagens (WT), CNR1 k/o e CNR2 k/o com EMS.
( A–F ) Representação do gráfico de barras ( média ± DP ) de esplenócitos (SP) corados para células T totais CD3 +, células T auxiliares CD4 +, células T citotóxicas CD8 +, células B CD19 +, monócitos CD11b + e F4/80 …
Painel de controle representando células T de esplenócitos.
( A–G ) Esplenócitos corados para CD3 no eixo y e CD11b no eixo x entre camundongos selvagens (WT), CNR1 k/o e CNR2 k/o com condição semelhante à endometriose (EM), bem como CNR1 k/o e CNR2 k/o …
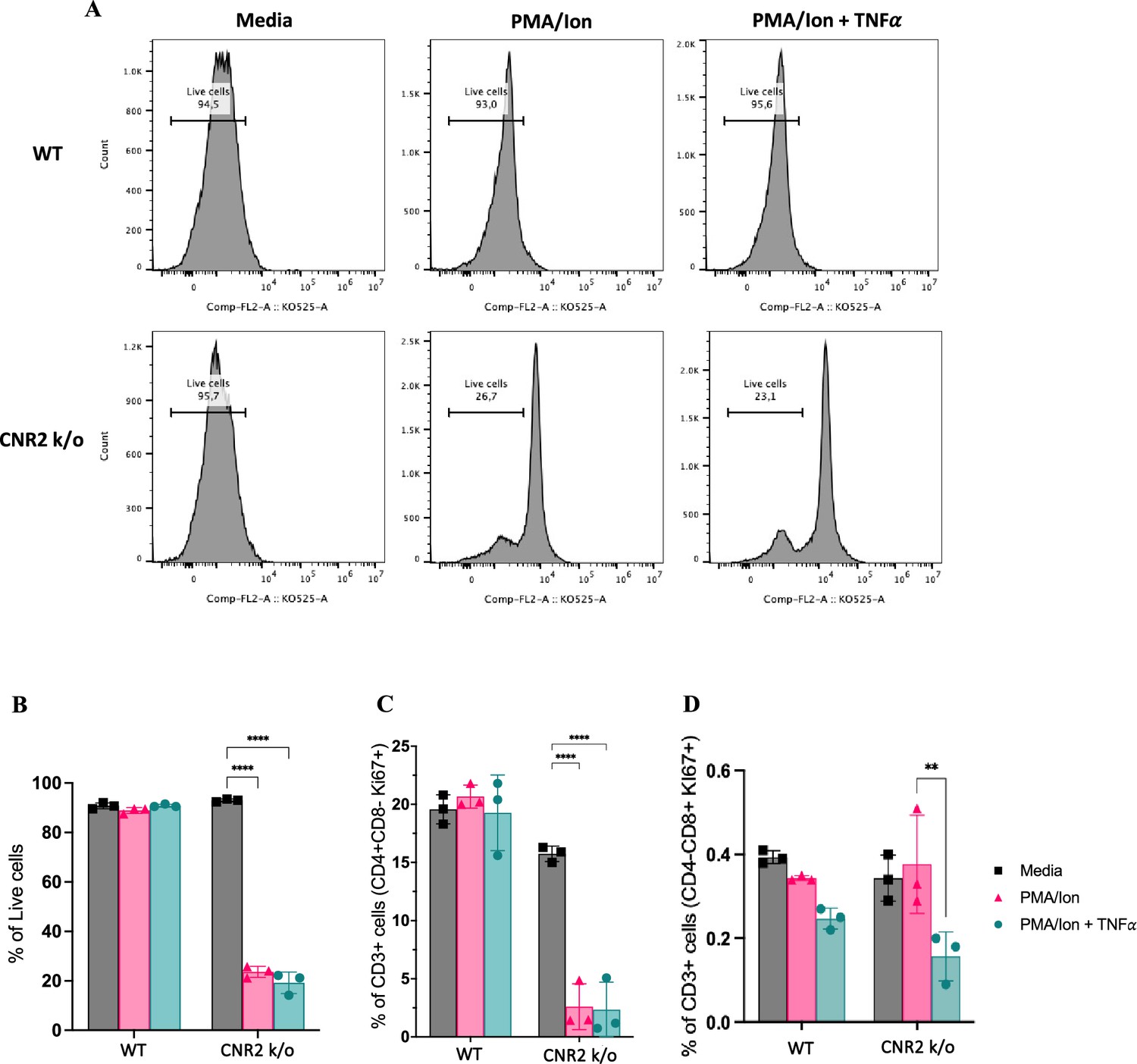
Células T de camundongos CNR2 k/o apresentam viabilidade prejudicada após ativação do TCR
Para validar as consequências funcionais da deficiência de CNR2 no comportamento das células T, conduzimos uma série de ensaios in vitro usando células T isoladas de esplenócitos de camundongos WT e CNR2 k/o naïve. Células T CD3+ foram ativadas de forma não específica, com ou sem coquetel de PMA/ionomicina, na presença ou ausência de fator de necrose tumoral alfa (TNFα) para criar um desafio inflamatório estéril.
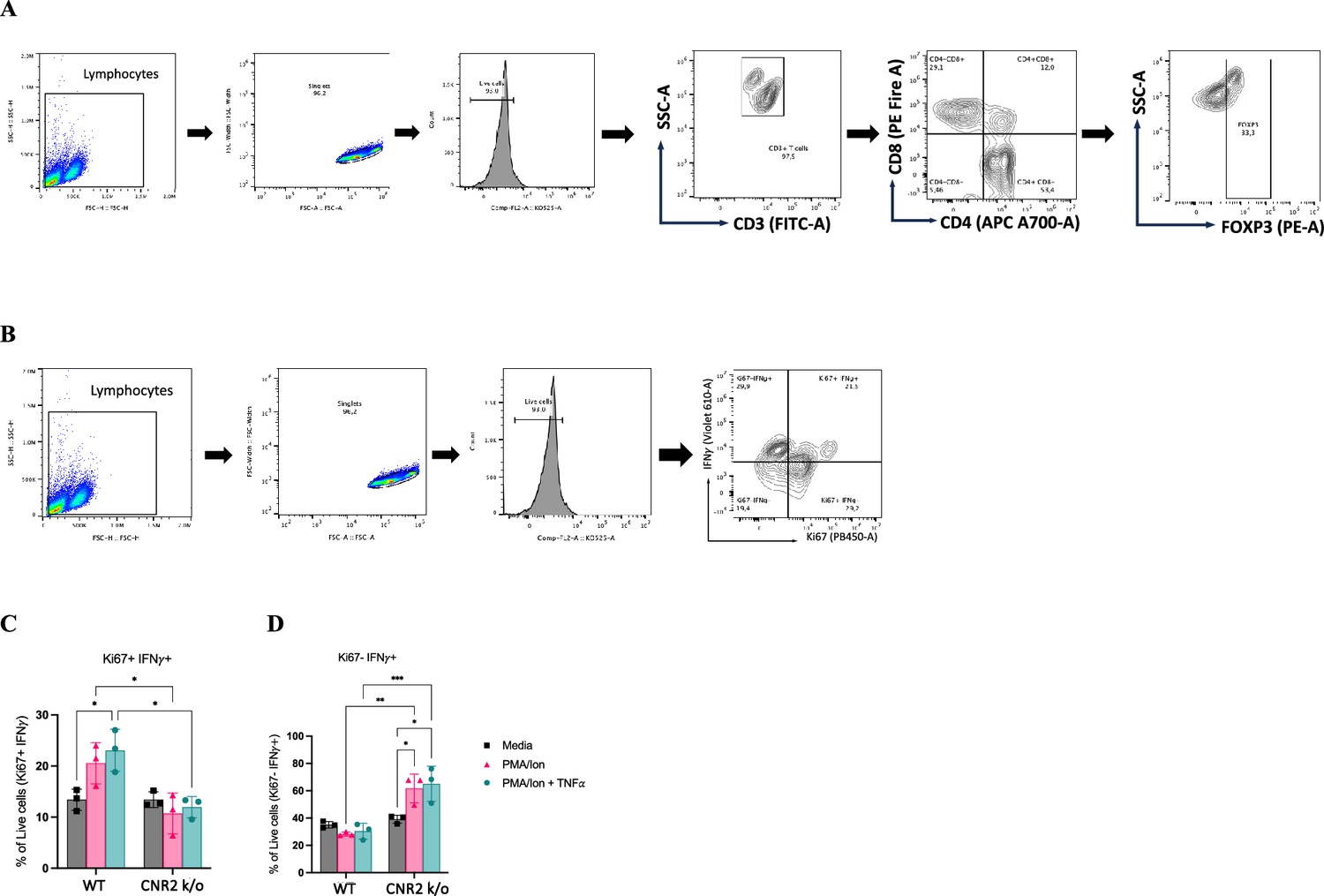
Observamos uma redução significativa na viabilidade de células T CD3 + totais de camundongos CNR2 k/o após ativação com PMA/ionomicina (estratégias de controle na Figura 5—suplemento da figura 1 ) em comparação aos controles de meio ( Figura 5A e B ). Em contraste, células T CD3 + WT ativadas com PMA/ionomicina, com ou sem TNFα, não exibiram nenhuma diferença significativa na viabilidade quando comparadas ao controle de meio ( Figura 5A e B ). Essa observação se alinha com nossas descobertas in vivo, nas quais células T CD3 + de camundongos CNR2 k/o com EM exibiram uma redução significativa na população esplênica e PF, mas não nos camundongos operados por SHAM, enfatizando a natureza específica de EM desse efeito. Além disso, a viabilidade geral reduzida de células T CD3 + de camundongos CNR2 k/o após ativação de PMA/ionomicina levou a uma diminuição na proliferação de células T CD4 + ( Figura 5C ), mas não de células T CD8 + ( Figura 5D ). No entanto, nossos resultados indicaram que, embora as células T deficientes em CNR2 apresentassem viabilidade reduzida após ativação, elas exibiram níveis mais altos de produção de IFNγ em comparação com células T CD3 + de camundongos WT, sugerindo sua competência funcional ( Figura 5—figura suplementar 1 ). Essas descobertas lançam luz sobre o papel intrincado do CNR2 na modulação das respostas das células T, com potenciais implicações para a desregulação imunológica.
Validação in vitro da deficiência de CNR2 na viabilidade e funcionalidade das células T CD3 em condições representativas do microambiente da lesão de endometriose (EM).
( A ) Representação da porcentagem de células T CD3 + totais vivas de camundongos selvagens (WT) e CNR2 k/o ativados com ou sem coquetel PMA/Ionomicina na presença ou ausência de necrose tumoral …
Ensaio funcional in vitro e avaliação por citometria de fluxo de células T CD3 ativadas de camundongos selvagens (WT) e CNR2 k/o ingênuos.
( A, B ) Estratégias de citometria de fluxo para células T CD3 + para identificar os diferentes fenótipos e estados funcionais, respectivamente. ( C ) Gráficos de barras representando a porcentagem de células vivas positivas que …
Análise espacial de células de lesões EM de camundongos WT, CNR1 k/o e CNR2 k/o revela populações de células imunes alteradas
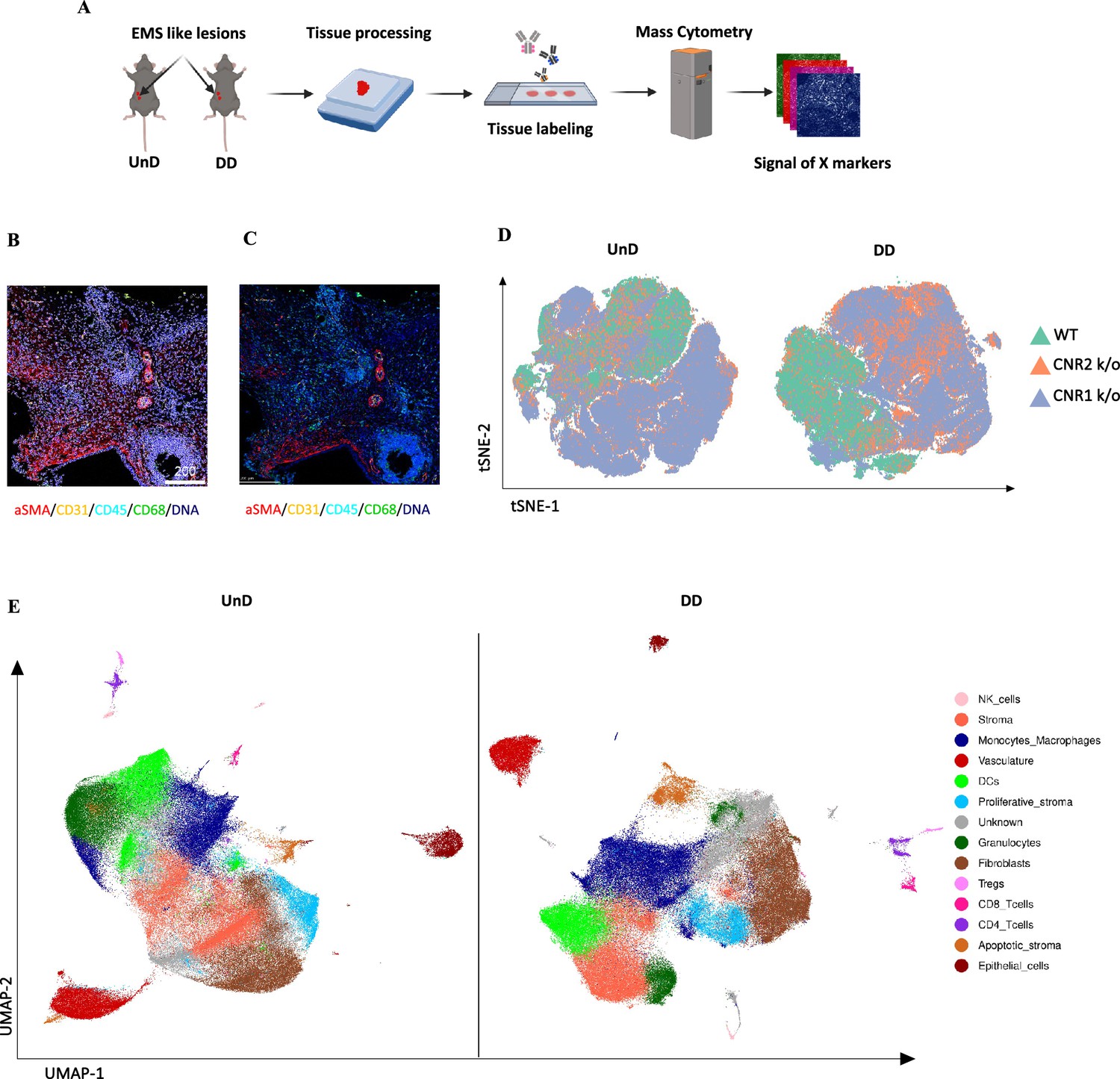
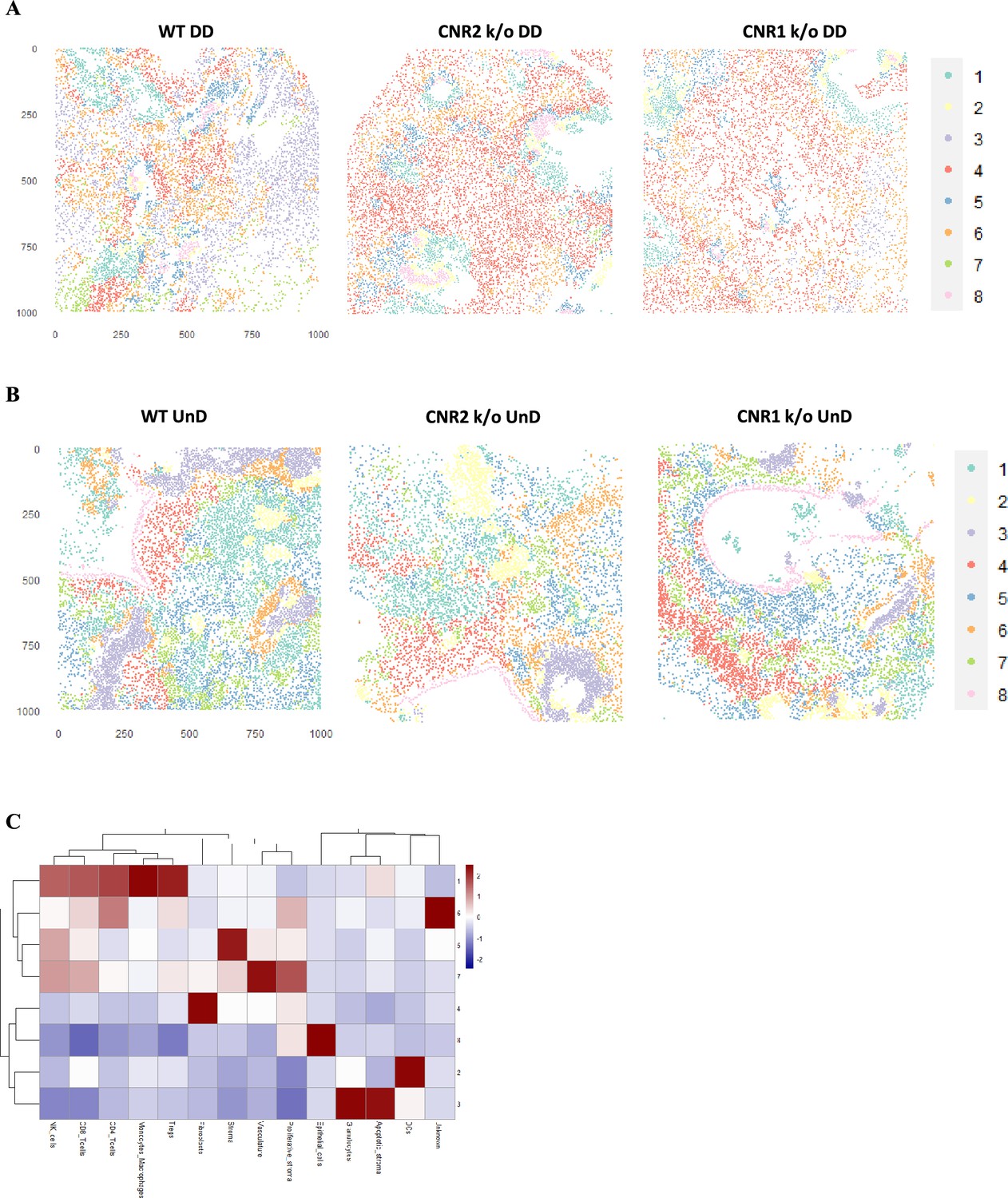
Para obter uma visão abrangente da distribuição espacial de células imunes e tipos de células estromais dentro da arquitetura de lesões EM de lesões UnD e DD de camundongos WT, CNR1 k/o e CNR2 k/o, empregamos a análise IMC. Essa abordagem teve como objetivo elucidar o impacto da ausência de CNR1 e CNR2 na composição e organização celular de lesões EM em camundongos. A representação esquemática do procedimento IMC ( Figura 6A ) descreve as etapas envolvidas nessa análise. Após a aquisição de duas regiões de interesse (ROI) por seção (com base na coloração H&E), a segmentação de célula única ( Figura 6B ) e a subsequente avaliação da qualidade da segmentação ( Figura 6C ) foram realizadas. Após a correção do efeito de lote das amostras, a redução da dimensionalidade não linear do tipo de amostra revelou um padrão de expressão distinto de células imunes e marcadores de estado celular entre lesões UnD e DD, bem como diferenças entre os genótipos ( Figura 6D ). Após a fenotipagem e marcação não supervisionadas dos diferentes tipos celulares, a redução da dimensionalidade por aproximação e projeção de coletor uniforme (UMAP) evidenciou ainda mais as principais diferenças na composição celular entre as lesões UnD e DD ( Figura 6E ). No geral, a expressão combinada dos tipos celulares das lesões DD de todos os três genótipos exibiu aumento dos compartimentos estromais, diminuição das células epiteliais e aumento da infiltração de macrófagos em comparação com a expressão dos tipos celulares das lesões UnD. Imagens representativas ilustram a distribuição dos diferentes tipos celulares com base no agrupamento e marcação não supervisionados ( Figura 7A ).
Perfil espacial de citometria de massa por imagem da distribuição de células imunes e padrões celulares em lesões de endometriose (EM) em camundongos CNR1 k/o, CNR2 k/o e tipo selvagem (WT).
( A ) O fluxo de trabalho de coleta e análise de dados de citometria de massa de imagem (IMC) descreve as etapas envolvidas na obtenção de insights abrangentes sobre a distribuição espacial de células imunes e ...
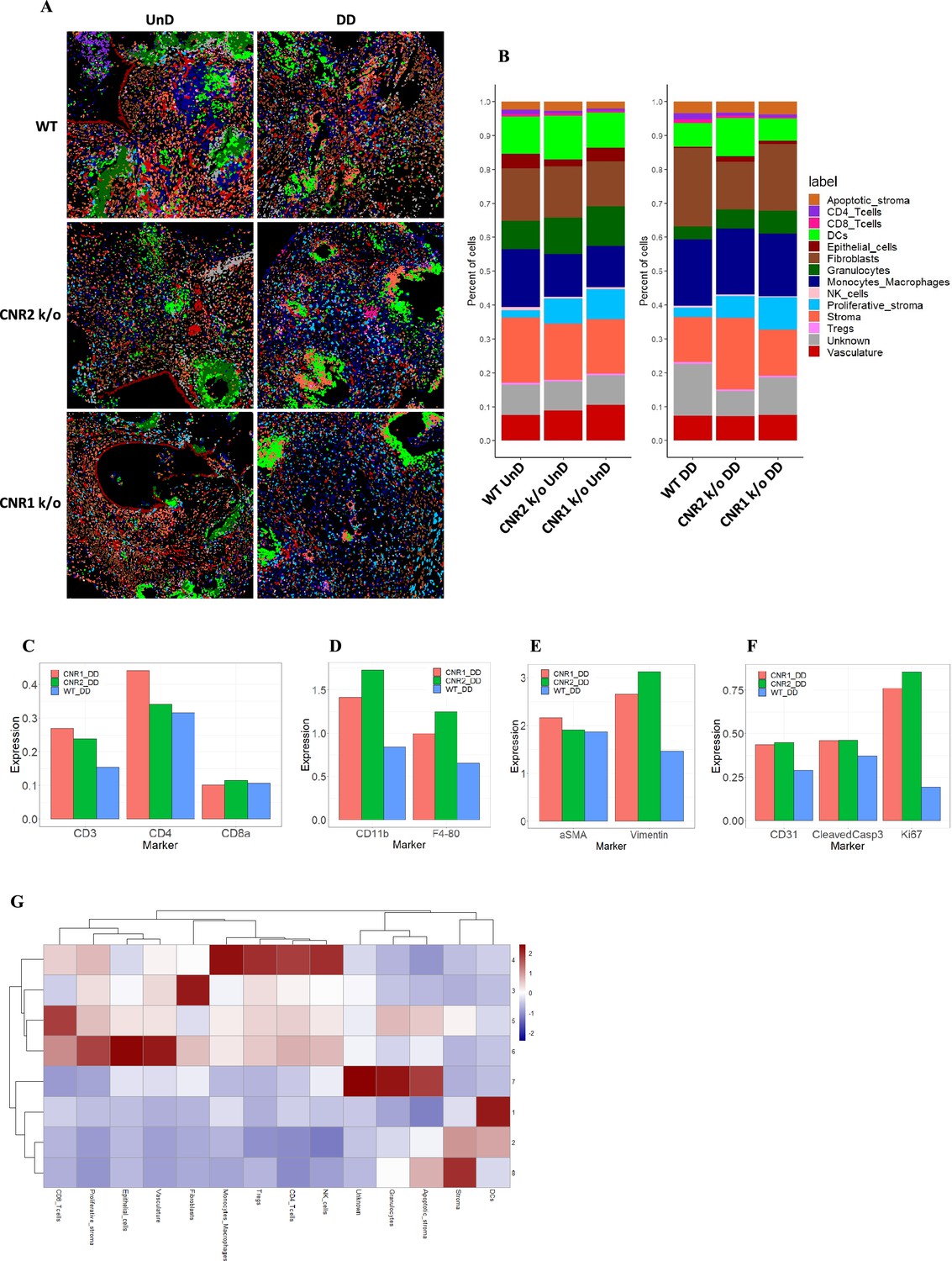
A citometria de massa por imagem revelou composição celular alterada e vizinhanças em lesões de endometriose (EM) de camundongos CNR1 k/o, CNR2 k/o e do tipo selvagem (WT).
( A ) Imagem representativa mostrando a distribuição de diferentes tipos de células dentro de lesões EM (n = 2–3 por tipo de tecido) com base em agrupamento e rotulagem não supervisionados. ( B ) Os gráficos de barras empilhadas revelam o …
Imagem de citometria de massa (IMC) e vizinhanças celulares (CN) em lesões não decidualizadas (UnD), EMS de camundongos CNR1 k/o, CNR2 k/o e tipo selvagem (WT).
( A, B ) Imagem representativa de oito tipos distintos de células da análise da vizinhança celular (CN) de lesões decidualizadas (DD) e UnD de camundongos WT, CNR1 k/o e CNR2 k/o, respectivamente. ( C ) Mapa de calor …
Análises adicionais da distribuição do tipo celular ( Figura 7B ) por meio de gráficos de barras revelaram diversas diferenças. Embora não significativas, devido ao número relativamente baixo de réplicas biológicas, a expressão de células T foi aumentada (células T auxiliares CD4 + e células T citotóxicas CD8 +) em lesões UnD e DD de camundongos CNR1 k/o e CNR2 k/o em comparação com WT ( Figura 7C ). Essa observação destaca que as células T residentes não são impactadas na ausência de CNR1 e CNR2 dentro do meio endometriótico. Curiosamente, em lesões de EM de CNR1 k/o e CNR2 k/o, vimos expressão significativamente elevada de monócitos/macrófagos ( Figura 7D ), células estromais ( Figura 7E ) e características de EM, como proliferação e vascularização ( Figura 7F ), demonstrando um microambiente alterado na ausência desses receptores. Para compreender as interações célula-célula e suas implicações, conduzimos uma análise de vizinhança celular (CN). Esta abordagem agrupou células com base em informações dentro de sua vizinhança espacial direta e identificou relações espaciais complexas entre diversos tipos de células dentro do microambiente da lesão. Esta análise revelou padrões de agrupamento distintos entre diferentes tipos de células dentro da arquitetura da lesão das lesões DD de camundongos WT, CNR1 k/o e CNR2 k/o ( Figura 7—suplemento da figura 1 ). As células imunes se agruparam predominantemente no NC 4, enquanto outros tipos de células (estroma, células epiteliais e vasculatura) exibiram padrões de agrupamento distintos entre os NC 6, 3 e 8 nas lesões DD ( Figura 7G ). Embora a maioria dos tipos de células imunes se agrupassem nas lesões UnD, os tipos de células da arquitetura da lesão se agruparam distintamente em comparação com as lesões DD ( Figura 7—suplemento da figura 1 ). Este agrupamento enfatiza a interação entre as células imunes e os componentes celulares mais amplos das lesões. Em resumo, nossa investigação abrangente revelou relações espaciais complexas entre células imunes e diversos tipos de células dentro de lesões EM em camundongos. As alterações observadas na expressão das células T, juntamente com a dinâmica estromal, nas lesões CNR1 k/o e CNR2 k/o ressaltam o papel fundamental desses receptores na formação do microambiente endometriótico.
Discussão
Evidências emergentes de nossas descobertas relatadas anteriormente e outras implicaram a desregulação do ECS, compreendendo receptores canônicos CNR1 e CNR2 junto com seus ligantes EC, na fisiopatologia do EM ( Lingegowda et al., 2021b ; Sanchez et al., 2016 ). O ECS está envolvido em vários processos fisiológicos, incluindo (mas não se limitando a) percepção da dor, regulação imunológica e funções reprodutivas Rodríguez de Fonseca et al., 2005 ; CNR1 e CNR2 são expressos em células imunes, tecidos nervosos e servem como reguladores críticos de processos reprodutivos, incluindo decidualização e implantação de embriões ( Di Blasio et al., 2012 ; Walker et al., 2019 ). Especula-se que a etiologia do EM seja decorrente da decidualização defeituosa e da menstruação retrógrada dos fragmentos endometriais, combinadas com a desregulação do SEC ( Lingegowda et al., 2022 ; Sampson, 1927 ; Maia et al., 2020 ). Embora seja plausível que a função diminuída do SEC influencie o início, a progressão e a experiência de dor intensa da lesão do EM, não está claro se a disfunção do SEC contribui ativamente para a patogênese do EM ou se representa uma consequência secundária de alterações que ocorrem no tecido endometrial refluído, levando ao estabelecimento de lesões do EM.
Mantendo esse dogma central em mente e para fornecer insights sobre os eventos iniciais da patogênese do EM, induzimos o EM em camundongos CNR1 k/o e CNR2 k/o utilizando fragmentos endometriais uterinos singênicos, DD e UnD. A ausência de CNR1 e CNR2 não influenciou os níveis sistêmicos de ligantes do ECS, mas o microambiente da lesão apresentou alterações significativas nos níveis de OEA e PEA, sugerindo uma resposta específica do tecido.
Um aspecto intrigante do envolvimento do ECS no EM é seu papel na decidualização, um processo essencial para a receptividade uterina à implantação do embrião e à gravidez bem-sucedida, que também pode contribuir para o estabelecimento do EM ( Correa et al., 2016 ). Embora tanto o CNR1 quanto o CNR2 sejam ativos na decidualização, o CNR1 pode ter um papel mais proeminente. A ausência de CNR1 e CNR2 mostra decidualização comprometida em camundongos de maneira dependente de CNR1 validada por meio de estudos in vitro ( Li et al., 2020 ). Da mesma forma, em nosso estudo, lesões de EM (tanto UnD quanto DD) de camundongos sem CNR1 mostraram significativamente mais DEGs (2088), em comparação com CNR2 (287) e WT (2). Genes essenciais para a decidualização, como Igfbp2 , Bmp3 , Ptgdr , Wnt7a e Esr1 foram regulados negativamente nas lesões DD e EM de camundongos CNR1 k/o em comparação com suas contrapartes UnD. Isso reforça ainda mais o papel do CNR1 no microambiente uterino e da lesão EM, incluindo seu papel na resposta de decidualização. Além disso, a interação entre CNR1 e CNR2 é importante, pois o CNR2 contribui para a imunomodulação, que é um processo-chave durante a decidualização ( Turcotte et al., 2016 ; Taylor et al., 2010 ). Embora a menstruação retrógrada seja considerada o principal mecanismo pelo qual os fragmentos endometriais atingem a cavidade peritoneal e se implantam para formar lesões EM, os próprios detritos menstruais retrógrados não sofrem decidualização. Entretanto, algumas lesões de EM na cavidade peritoneal, sob a influência de estrogênio e progesterona, podem sofrer decidualização, pois as lesões podem existir em diferentes estágios evolutivos, desde lesões vermelhas ativas até lesões brancas quiescentes ( Leone Roberti Maggiore et al., 2016 ).
Dada a complexidade da sinalização do SEC e dos mecanismos compensatórios, focamos nossa investigação no aspecto da desregulação imunológica da fisiopatologia do EM. Nossas descobertas sugerem que a dinâmica alterada do SEC durante a decidualização interrompe a sinalização do SEC, levando à desregulação das respostas imunológicas e ao comportamento celular aberrante. De fato, a disfunção imunológica é uma característica do EM ( Symons et al., 2018 ; Ahn et al., 2015 ), e o SEC pode desempenhar um papel crucial na formação das respostas imunológicas, particularmente por meio de seu impacto na função das células T, embora não haja evidências claras. Alterações nas populações e funções das células T têm sido associadas à progressão do EM, sugerindo seu papel vital na patogênese e manutenção do EM ( Tanaka et al., 2017 ; Kyama et al., 2003 ). Nossa análise de citometria de fluxo revelou alterações significativas nas populações de células imunes em camundongos portadores de lesões de EM, com uma notável ausência de células T CD3+, células T auxiliares CD4+ e células T citotóxicas CD8+ especificamente em camundongos CNR2 k/o. Estudos mecanísticos in vitro confirmaram ainda uma resposta aberrante de células T em camundongos CNR2 k/o, pois com a ativação e estimulação do receptor de células T houve diminuição da viabilidade. Combinados, nossos achados mostram que o CNR2 é crítico na sobrevivência de células T após a ativação do TCR com padrões moleculares associados a patógenos (PAMPs)/padrões moleculares associados a perigos (DAMPs) ou sinais mediados por antígenos. Esses achados também lançam luz sobre um papel anteriormente não reconhecido do CNR2 na disfunção imune adaptativa associada ao EM, dado o papel crítico das células T na vigilância e regulação imunológica. Além disso, a especulação de que o EM seja uma causa da disfunção do ECS pode ser importante, uma vez que o CNR2 foi encontrado reduzido nas lesões de pacientes com EM, conforme demonstrado por nosso estudo anterior ( Lingegowda et al., 2021b ). Além disso, o sequenciamento de RNA em massa reforça a descoberta de desregulação de genes relacionados a células T em lesões de EM de CNR2 k/o. A regulação negativa observada de genes relacionados a células T, como Cd3e , Cd3g , Gata3 e Ctla4 , é consistente com a diminuição das populações de células T CD3+ e destaca a relevância de CNR2 nas respostas imunes mediadas por células T no microambiente endometriótico.
A análise da via KEGG de genes relacionados ao sistema imunológico diferencialmente expressos em lesões de CNR2 k/o DD revelou ainda que as vias de diferenciação de células Th1, Th2 e Th17 foram impactadas, e a literatura anterior confirma a desregulação dessas vias na fisiopatologia do EM ( Chang et al., 2023 ; Andreoli et al., 2011 ). Além disso, nosso ensaio funcional in vitro, mostrando que as células T auxiliares CD4+ foram afetadas (proliferação) mais do que o subconjunto de células T citotóxicas CD8+, contribui ainda mais para o comportamento específico do subconjunto de CNR2. Nossos resultados fornecem um elo perdido entre o SEC e o funcionamento do sistema imunológico durante o EM.
Para obter insights sobre as características iniciais de iniciação e estabelecimento de lesões, conduzimos uma análise IMC de lesões EM em genótipos e diferentes tipos de lesão (DD e UnD). Lesões DD de camundongos CNR1 e CNR2 k/o mostraram maior célula T residindo nas lesões com aumento de compartimentos estromais e população de monócitos/macrófagos em comparação com lesões WT. As células estromais contribuem para o desenvolvimento inicial de lesões EM promovendo inflamação, angiogênese, fibrose e modulação imunológica ( McKinnon et al., 2022 ; Queckbörner et al., 2020 ; Zondervan et al., 2018 ). Além disso, a interação entre macrófagos e células estromais é importante no EM, com o inflamossomo NLRP3 desempenhando um papel no desenvolvimento da lesão ( Guo et al., 2021 ). Estudos têm implicado macrófagos no crescimento de lesões de EM, onde eles apoiam a angiogênese (formação de vasos sanguíneos) produzindo fatores pró-angiogênicos, como o fator de crescimento endotelial vascular (VEGF) ( Ding et al., 2012 ; Sun et al., 2019 ; McLaren et al., 1996 ). Dado o aumento combinado na proliferação, marcadores endoteliais e monócitos/macrófagos em lesões de EM de camundongos potencialmente indica que eles poderiam modular o microambiente inicial da lesão no caso de desregulação de CNR1 e CNR2. Com base na proporção desses macrófagos para certos fenótipos, como M1 ou M2, ditaria o desenvolvimento da lesão e a progressão subsequente. Mais estudos são necessários para desvendar as interações moleculares de CNR1 e CNR2 com subconjuntos específicos de células imunes em um microambiente complexo de lesão de EM e determinar como isso contribui para o estabelecimento do suprimento sanguíneo e a sobrevivência da lesão.
A desregulação do SEC, como evidenciado pela expressão alterada de CNR1 e CNR2 em nosso modelo murino de EM, parece ter consequências de longo alcance no cenário celular e molecular das lesões endometrióticas. Como o CNR1 é amplamente expresso no sistema nervoso central, onde regula a percepção da dor, o humor e outros processos neurológicos, a desregulação da sinalização do CNR1 pode impactar a inervação sensorial e as respostas à dor associadas ao EM ( Rodríguez de Fonseca et al., 2005 ). Além disso, o CNR1 demonstrou modular a função das células imunes, incluindo a produção de citocinas pró-inflamatórias Klein, 2005 . A perda da sinalização do CNR1 em células imunes que infiltram as lesões endometrióticas provavelmente contribui para os perfis imunes alterados observados em nosso estudo. Em contraste, o CNR2 é predominantemente expresso em células imunes, como macrófagos, linfócitos e células natural killer ( Turcotte et al., 2016 ). Este receptor desempenha um papel crucial na regulação das respostas imunes, incluindo a modulação da produção de citocinas, migração celular e proliferação. A redução significativa de células T CD3+ na cavidade peritoneal e no baço de camundongos com CNR2 knockout sugere que a perda de CNR2 pode ter um impacto profundo na homeostase e na função das células T de maneira dependente de ME, o que não ocorreu em camundongos submetidos a cirurgia simulada. Esse comprometimento na função das células T tem consequências diretas na resposta imune adaptativa, como evidenciado pelos perfis de expressão gênica alterados e pelas vias relacionadas à função imune em nossa análise transcriptômica. Contudo, não conduzimos experimentos mecanísticos específicos para desvendar a interação entre a resposta imune e o desenvolvimento de lesões de endometriose em nossos sistemas modelo.
Várias limitações devem ser reconhecidas em nosso estudo. Primeiramente, compreender os aspectos homeostáticos do SEC, tanto com a presença de CNR1 quanto sem a presença de CNR1 e CNR2, continua sendo um desafio complexo. Embora nossos modelos globais de camundongos k/o forneçam insights valiosos, pesquisas futuras utilizando k/o direcionado específico para o útero podem oferecer uma compreensão mais precisa de suas contribuições para a fisiologia uterina e implicações adicionais no estabelecimento de EM. O uso de modelos de camundongos para estudar EM tem limitações inerentes devido às diferenças entre as espécies e à incapacidade de recapitular completamente a doença humana. As complexidades moleculares e celulares associadas aos tecidos endometriais DD e UnD, bem como o momento da indução de EM nesses camundongos, podem não refletir perfeitamente a condição humana. Não foram observadas alterações basais significativas no perfil imunológico entre os camundongos k/o CNR1 e CNR2 e os camundongos WT sem indução de EM. No entanto, o ambiente uterino não foi avaliado para compreender o perfil imunológico basal entre os camundongos k/o e WT. Essas limitações enfatizam a necessidade de investigações futuras para aumentar a relevância translacional de nossas descobertas e aprofundar nossa compreensão da interação complexa entre ECS, decidualização e patogênese do EM.
Em conclusão, nosso estudo oferece evidências do envolvimento da desregulação de CNR1 e CNR2 na patogênese do EM. Por meio de uma análise integrativa de perfis transcriptômicos, dinâmica de células imunes e relações espaciais em lesões de EM de camundongos, desvendamos as intrincadas interações entre o SEC, as respostas imunes e as alterações celulares no EM. Ao identificar mecanismos potenciais pelos quais a interrupção do SEC pode impactar o EM, nossa pesquisa fornece uma base para o desenvolvimento de terapias direcionadas que abordem a influência do SEC no EM. Essas descobertas ampliarão nossa compreensão do EM e levarão a estratégias terapêuticas inovadoras para o manejo desse distúrbio complexo.
Métodos
Animais
Os experimentos descritos neste trabalho foram aprovados pelo Comitê Institucional de Cuidados com Animais da Queen's University, de acordo com as diretrizes fornecidas pelo Conselho Canadense de Cuidados com Animais para o protocolo número 2021–2228. Todos os animais foram designados aleatoriamente para os procedimentos cirúrgicos. Todos os estudos foram realizados usando camundongos fêmeas adultas com idades entre 7 e 10 semanas. Camundongos reprodutores machos e fêmeas CNR1 k/o (B6.129P2(C)- Cnr1 tm1.1Ltz/J) e CNR2 k/o (B6.129P2- Cnr2 tm1Dgen/J) foram obtidos do Jackson Laboratory (Bar Harbor, EUA) e alojados no biotério da Queen's University. Camundongos fêmeas experimentais CNR1 k/o e CNR2 k/o foram obtidos por cruzamento trio com seus respectivos machos homozigotos k/o. Camundongos fêmeas controle C57BL/6 j (WT) e camundongos machos vasectomizados com 7 a 10 semanas foram obtidos do Jackson Laboratory. Todos os camundongos reprodutores foram alojados em gaiolas de reprodução padrão em uma instalação de barreira, e os animais experimentais foram alojados em uma área de contenção convencional. Os animais foram alojados em temperatura constante (22 ± 1 °C) e umidade relativa (50%), com um ciclo claro-escuro de 12:12 h (luz acesa das 7h às 19h). Comida e água estavam disponíveis ad libitum. Todos os animais experimentais foram aclimatados na instalação de contenção convencional por 1 semana antes do início dos experimentos.
Descidualização in vivo
Neste estudo, usamos um modelo murino singênico modificado de EM, onde os fragmentos doadores foram obtidos de cornos uterinos decidualizados artificialmente. O método de decidualização artificial usado neste estudo foi previamente estabelecido e utilizado por vários estudos de pesquisa ( Cai et al., 2022 ; Benson et al., 1996 ; Liu et al., 2022 ). Para induzir artificialmente a decidualização, camundongos fêmeas foram autorizados a acasalar com camundongos machos vasectomizados para induzir pseudogestação. Após o 4º dia de pseudogestação, camundongos fêmeas foram submetidos à laparotomia para receber uma injeção de 30 μL de óleo de semente de gergelim (S3547, Sigma, EUA), intraluminalmente em um corno uterino para induzir DD. O corno contralateral não injetado serviu como um controle UnD. Após a injeção de óleo de gergelim, os animais permaneceram em repouso por 4 dias, após os quais a DD foi induzida com sucesso em um corno uterino, conforme mostrado na Figura 1A . Esses cornos uterinos foram utilizados como fragmentos doadores para induzir EM em camundongos receptores de seus respectivos genótipos. A Figura 1B mostra as imagens representativas das lesões de EM 7 dias após a indução cirúrgica.
Modelo de camundongo de EM
O EM foi induzido cirurgicamente conforme descrito anteriormente ( Lingegowda et al., 2021b ; Lingegowda et al., 2021a ). Dois grupos independentes (DD e UnD) por genótipo foram usados neste estudo (n = 8–16). Resumidamente, os cornos uterinos DD e UnD dos camundongos doadores foram colhidos e os cornos uterinos foram dissecados longitudinalmente para revelar o endométrio. Fragmentos uterinos foram obtidos usando um punch de biópsia epidérmica de 3,0 mm (33–32, Integra Miltex, EUA). Os camundongos receptores foram anestesiados sob anestesia com vaporizador de isoflurano a 3,5% para fazer uma incisão na linha média no abdômen (n = 8–16) e dois fragmentos uterinos DD ou UnD de 3,0 mm foram implantados na parede peritoneal interna direita usando uma cola de ligação de tecidos de grau veterinário (1469 SB, 3 M, EUA). Os grupos controle WT, CNR1 k/o e CNR2 k/o (n=4) foram submetidos a cirurgia simulada com uma incisão na linha média do abdômen sem implantação de fragmentos uterinos. Os camundongos foram sacrificados 7 dias após a cirurgia de indução de EM, uma vez que o foco do estudo estava no ponto de tempo anterior ao início da EM após a indução. Com base em estudos anteriores, 7 dias após a indução de EM parece ser a fase logarítmica do reparo tecidual e da resposta imune ( Symons et al., 2020 ). O sangue foi colhido por punção cardíaca para avaliar os ligantes de EC. O fluido peritoneal (PF) foi coletado pela injeção de 3 ml de solução salina tamponada com fosfato (PBS) gelada na cavidade peritoneal. Os baços foram coletados em meio RPMI gelado (11875093, ThermoFisher, Canadá) antes do processamento para obter suspensão de células únicas. As lesões de EM foram congeladas rapidamente em nitrogênio líquido e armazenadas a –80℃ ou processadas usando paraformaldeído 4% durante a noite (12–20 horas), mantidas a 4 °C em etanol 70% e então incluídas em parafina.
Extração de lipídios e espectrometria de massa direcionada
O plasma não foi diluído e ~10 mg de tecido por amostra foram homogeneizados com tampão RIPA (89900, Thermo Fisher, Canadá) com 1:100 de coquetel inibidor de protease (535140–1 ML, Sigma, Canadá) para obter lisados de tecido. Tanto o plasma quanto os lisados de tecido obtidos foram submetidos individualmente à extração em fase sólida (SPE). Os padrões internos, deuterados e não deuterados, usados no estudo para avaliar os ligantes ECS foram adquiridos da Cayman Chemicals, EUA ( arquivo suplementar 1a ). Alíquotas de 100 µL de plasma e lisados de tecido foram adicionadas a uma placa de precipitação de proteínas (CE0-7565-R, Phenomenex, EUA) juntamente com 200 µL de acetonitrila fria contendo os padrões internos deuterados. O filtrado foi diluído com 500 μL de água e submetido à extração por SPE em uma placa Oasis HLB de 96 poços (WAT058951, Waters, Canadá). As amostras foram lavadas com 60% de metanol antes da eluição com acetonitrila. O eluato foi seco, reconstituído em 100 μL da fase móvel A e analisado por cromatografia líquida com espectrometria de massas (LC-MS, conforme descrito no arquivo suplementar 1b ). As concentrações endógenas dos quatro compostos no plasma humano foram calculadas por adição padrão.
Citometria de fluxo
Suspensões unicelulares foram preparadas a partir de baços murinos por dissociação mecânica, lise de hemácias e centrifugação. Células esplênicas e células de PF foram ressuspensas em um tampão de coloração (PBS com 2% de soro fetal bovino) a uma concentração de 0,5 × 10^6 células/mL. Todos os anticorpos utilizados para análises de citometria de fluxo foram adquiridos da BioLegend, EUA, salvo indicação em contrário. Os anticorpos incluíram CD45-FITC (103107), CD3-BV510 (100234), CD4-BV785 (100551), CD8-BV605 (100744), CD11b-AF700 (101222), F4/80-PE/Cy7 (123114), NK1.1-APC/Cy7 (108724) e CD19-PE/Dazzle 594 (115554). A coloração foi realizada seguindo as recomendações do fabricante. Para cada amostra, 50 μL do coquetel de anticorpos foram adicionados a 50 μL de suspensão celular em placas de 96 poços. A mistura foi incubada a 4 °C por 20 minutos no escuro, juntamente com um anticorpo bloqueador de Fc anti-CD16/32 (101319). Após a incubação, as células foram lavadas duas vezes com tampão de coloração e centrifugadas antes da fixação com tampão de fixação (00-8222/49, Thermo Fisher, Canadá). A análise por citometria de fluxo foi realizada utilizando um citômetro de fluxo Beckman Coulter CytoFlex S. Os controles de compensação foram estabelecidos utilizando células coradas com anticorpo único. Os controles de isótipo forneceram níveis basais de coloração não específica e as populações celulares foram definidas utilizando o controle de fluorescência menos um (FMO). A análise de dados utilizou o software FlowJo (v 10.9; FlowJo, EUA), bem como o kit de ferramentas computacionais SPECTRE (v 1.0) em R (v 4.2.3) para obter gráficos de incorporação estocástica de vizinhos (t-SNE) distribuídos em t com base nos clusters flowSOM não supervisionados gerados pela expressão do marcador.
Ensaio funcional de células T in vitro
Células T CD3+ totais foram isoladas de esplenócitos de camundongos WT e CNR2 k/o naïve usando um kit magnético de seleção negativa (19851 A, StemCell, Canadá) seguindo as instruções do fabricante. Todas as proteínas e compostos recombinantes foram adquiridos da Biolegend, EUA, a menos que indicado de outra forma. Posteriormente, 250.000 células T por poço foram semeadas em uma placa de 96 poços revestida com anti-CD3 de camundongo [(2 μg/ml), (100340)]. Meio RPMI-1640 suplementado com rmIL-2 [(10 ng/ml), (575404)], anti-CD28 de camundongo [(5 μg/ml), (102116)], 10% de soro fetal bovino, β-mercaptoetanol (50 μM) e penicilina/estreptomicina (100 U/ml) foi usado como meio de crescimento. As células T foram então ativadas de forma não específica, com ou sem o coquetel de ativação celular composto por 12-miristato 13-acetato de Phorbol (PMA) e ionomicina [(50 ng/ml de PMA e 1 μM de ionomicina), (423301)] na presença e ausência de TNFα [(100 ng/ml), (410-MT-010/C, R&D Systems, EUA)] para simular um desafio inflamatório estéril. Após um período de incubação de 48 horas, brefeldina A [(10 μg/ml), (11861, Cayman Chemicals, EUA)] foi introduzida nas células para medir os níveis intracelulares de interferon-gama (IFNγ) no ponto de tempo de 42 horas. A análise por citometria de fluxo foi conduzida utilizando um painel de marcadores, adquiridos da Biolegend, EUA, salvo indicação em contrário, incluindo CD3e-FITC (100306), CD4-AF700 (100430), CD8-PE/fire700 (100792), Ki67-PB (151223), FoxP3-PE (126404), IFNγ-BV605 (505840) e Live/dead-K0525 (L304966, Thermo Fisher, Canadá), para avaliar vários subconjuntos de células T e sua viabilidade. A coloração e aquisição por citometria de fluxo foram realizadas conforme descrito acima, com a adição de tampão de permeabilização (00-8333-56, Thermo Fisher, Canadá) para a coloração de marcadores intracelulares (Ki-67 e IFNγ), de acordo com as instruções do fabricante. A análise dos dados foi realizada utilizando o software FlowJo e visualizada utilizando o GraphPad Prism (v 9.5.1).
Isolamento de RNA usando o mini kit RNeasy
Lesões de EM UnD e DD congeladas rapidamente de WT, CNR1 k/o e CNR2 k/o foram homogeneizadas, e o RNA foi isolado usando o RNeasy Mini Kit (74104, Qiagen, Canadá), de acordo com as instruções do fabricante. Resumidamente, tecidos de lesão de EM com ~20 mg foram colocados individualmente em tubos cerâmicos PowerBead (13113–50, Qiagen, Canadá) juntamente com tampão de lise. As lesões foram homogeneizadas usando um homogeneizador Bead Ruptor (Omni International, EUA) e os lisados foram extraídos após centrifugação a 10.000 RCF. O lisado foi misturado com etanol a 70%, adicionado à coluna de centrifugação RNeasy e, em seguida, centrifugado para ligar o RNA à coluna. A coluna de centrifugação foi lavada duas vezes, e o RNA foi isolado usando um tampão de eluição. A qualidade total do RNA foi medida usando o espectrofotômetro nanodrop e armazenada a –80°C antes do envio para a BGI Global (Boston, EUA) para análise de RNA em massa. A integridade do RNA foi determinada usando o sistema Agilent 4150 TapeStation System (Agilent, EUA) para controle de qualidade da amostra, e apenas amostras com número de qualidade de RNA ≥7 foram consideradas para preparação da biblioteca e sequenciamento posterior.
Preparação, sequenciamento e análise de biblioteca de RNA
A preparação da biblioteca começou com o enriquecimento do mRNA usando esferas de oligo dT, que capturam seletivamente moléculas de mRNA. Em seguida, o mRNA enriquecido foi fragmentado e o cDNA da primeira fita foi sintetizado usando primers N6 aleatórios, seguido pela síntese do cDNA da segunda fita usando trifosfato de desoxiuridina (dUTP). Após a síntese do cDNA, o reparo da extremidade foi realizado para gerar extremidades rombas e a adenilação 3' foi realizada para facilitar a ligação do adaptador. Os adaptadores foram ligados aos fragmentos de cDNA adenilados 3'. Para enriquecer a biblioteca de cDNA para sequenciamento, foi realizada a amplificação por PCR. Antes da amplificação, a fita marcada com dUTP foi especificamente degradada pela Uracil-DNA-Glicosilase (UDG). O cDNA da primeira fita restante foi então amplificado usando primers de PCR. Após a amplificação, a separação da fita simples foi obtida por desnaturação por calor. O DNA fita simples foi ciclizado usando um oligo splint e uma DNA ligase. A síntese de nanobolas de DNA foi realizada em moldes de DNA fita simples ciclizados. Esse processo facilitou a geração de clusters clonais de DNA, fornecendo o material necessário para o sequenciamento subsequente. O sequenciamento foi executado usando a plataforma DNBSEQ Technology. As bibliotecas de DNA preparadas foram carregadas no sequenciador DNBSEQ e sequenciadas a uma profundidade média de 30 milhões de leituras pareadas (2 × 100) por biblioteca. Os dados de sequenciamento foram filtrados com SOAPnuke, removendo leituras contendo o adaptador de sequenciamento; removendo leituras cuja razão de bases de baixa qualidade (qualidade de base menor ou igual a 15) é maior que 20% e removendo leituras cuja razão de bases desconhecida (base 'N') é maior que 5%. Em seguida, leituras limpas foram obtidas e armazenadas no formato FASTQ. As leituras limpas foram mapeadas para o genoma de referência do camundongo (NCBI: GRCm38.p6) usando HISAT2 (v2.0.4). A análise subsequente e a mineração de dados foram realizadas no sistema de mineração de dados Dr. Tom Multi-omics ( https://biosys.bgi.com ).
Citometria de massa por imagem: rotulagem
Um painel abrangente de anticorpos que identificam populações de células imunes inatas e adaptativas e tipos de células que são integrais ao microambiente da lesão EM ( arquivo suplementar 1c ) foi projetado e otimizado conforme descrito anteriormente ( McDowell et al., 2021 ; Sorin et al., 2023 ). As seções de tecido fixadas em formalina e embebidas em parafina (FFPE) (n = 2–3 por tipo de tecido) foram submetidas à desparafinização e recuperação de antígeno mediada por calor na plataforma de autocoloração Ventana Discovery Ultra (Roche Diagnostics, Canadá), seguindo as instruções abaixo. Inicialmente, as lâminas foram expostas a uma temperatura de 70 °C em uma solução EZ Prep pré-formulada (Roche Diagnostics, Canadá), seguida por uma incubação subsequente a 95 °C em solução Cell Conditioning 1 pré-formulada (Roche Diagnostics, Canadá). Em seguida, as lâminas foram lavadas em PBS 1x e, em seguida, expostas à solução Dako Serum-free Protein Block (Agilent, EUA) por 45 minutos à temperatura ambiente. Um coquetel de anticorpos contendo anticorpos conjugados a metais foi preparado usando o Diluente de Anticorpos Dako (Agilent, EUA) em diluições específicas. Os anticorpos primários contidos nesse coquetel foram aplicados às lâminas e deixados reagir durante a noite a 4°C, após o que as lâminas foram lavadas com Triton X-100 a 0,2% e PBS 1x. Para a etapa subsequente, um coquetel de anticorpos secundários contendo anticorpos antibiotina conjugados a metais foi criado no Diluente de Anticorpos Dako, em uma diluição predeterminada. As lâminas foram tratadas com esse coquetel antibiotina por 1 hora à temperatura ambiente e, em seguida, lavadas com Triton X-100 a 0,2% e PBS 1x. Para a contracoloração, as lâminas foram expostas ao Cell-ID Intercalator-Ir (Fluidigm, Canadá) diluído na proporção de 1:400 em PBS 1x por 30 minutos à temperatura ambiente. Após 5 minutos de enxágue com água destilada, as lâminas foram secas ao ar em preparação para a aquisição do IMC. O Hyperion Imaging System (Fluidigm, Canadá) foi utilizado para o processo de aquisição do IMC.
Citometria de massa por imagem: Análise de dados
Lesões (n = 2–3 por tipo de tecido) foram agrupadas com base na origem dos fragmentos uterinos (ou seja, UnD ou DD) de três genótipos diferentes (WT, CNR1 k/o e CNR2 k/o). Os métodos de análise de dados IMC empregados neste estudo seguem procedimentos estabelecidos conforme descrito no kit de ferramentas Steinbock (Spatial Experiment v 1. 12. 0) para pré-processamento de dados, segmentação de imagens e quantificação de objetos ( Windhager et al., 2021 ). A segmentação celular utilizou uma abordagem de aprendizado profundo descrita por Greenwald et al., 2022. Resumidamente, imagens de canal duplo foram geradas usando marcadores nucleares e citoplasmáticos, representando respectivos sinais. A ferramenta DeepCell com Mesmer, um algoritmo de segmentação de aprendizado profundo pré-treinado do TissueNet, foi usada para automatizar a geração de máscara celular, não exigindo entrada adicional do usuário. Dado que os dados do IMC foram adquiridos em lotes, realizamos correções de efeito de lote usando o algoritmo de harmonia conforme descrito ( Korsunsky et al., 2019 ). Isso envolveu agrupamento iterativo e correção de posições de células no espaço de análise de componentes principais (PCA). Posteriormente, o agrupamento PhenoGraph não supervisionado em R (v 4. 3. 2) foi usado para categorizar os tipos de células. Para isso, sinais incluindo αSMA, B220, CD19, β-catenina, CD3, CD4, CD8, CD11b, CD11c, CD31, CD68, E-caderina, MPO, pan-citoqueratina e vimentina foram utilizados, empregando um valor k de 60. Para verificar as interações celulares, imcRtools (v 1. 8. 0) e cytomapper (v 1. 14. 0) em R foram empregados para visualização. Um teste de permutação avaliou as interações com células vizinhas. Células vizinhas foram definidas como aquelas dentro de um raio de cinco pixels (5 µm), e a função buildSpatialGraph estabeleceu o número de vizinhos de um cluster interagindo com outro cluster. Um padrão de 1000 permutações foi definido. Cada iteração levou a uma pontuação de interação e ao cálculo do valor de p, e os resultados significativos (com risco alfa de 1%) foram representados em mapas de calor. Para delinear as vizinhanças celulares espaciais, janelas vizinhas foram computadas, representando as N células mais próximas de cada célula. Este processo seguiu protocolos anteriores. Empregando imcRtools, o agrupamento de vizinhanças celulares foi conduzido, levando à identificação de 8 vizinhanças celulares nas lesões.
Estatísticas
As análises estatísticas realizadas para comparar a concentração de ligantes de EC por LC-MS direcionado e a avaliação da população de células imunes por citometria de fluxo foram conduzidas utilizando o Prism GraphPad. Uma análise de variância (ANOVA) unidirecional foi realizada com o teste post-hoc de Holm-Sidak para determinar as diferenças específicas entre os grupos. Para o ensaio funcional de células T in vitro, a ANOVA bidirecional foi realizada utilizando o teste post-hoc de Tukey para comparar dentro e entre os grupos. O nível de significância foi estabelecido em α = 0,05. Os dados são apresentados como média ± desvio padrão (DP), salvo indicação em contrário.
Disponibilidade de dados
Os dados de sequenciamento de mRNA em massa são fornecidos nos arquivos suplementares (Dados Suplementares 4) e os dados de IMC gerados neste estudo foram depositados no Mendeley Data ( https://doi.org/10.17632/2ptns5yhzh.2 ). Todos os códigos utilizados no estudo atual são de artigos publicados anteriormente, conforme citados no texto. Os autores não relatam o código original.
Institutos Canadenses de Pesquisa em Saúde (394340)
Chandrakant Tayade
Os financiadores não tiveram nenhum papel no desenho do estudo, na coleta e interpretação de dados, ou na decisão de submeter o trabalho para publicação.
Agradecimentos
Agradecemos à Dra. Alexandra Furtos e Karine Gilbert, do Centro Regional de Espectrometria de Massas da Universidade de Montreal, pelo projeto e execução da avaliação por espectrometria de massas; ao Dr. Yuhong Wei e à plataforma de citometria de massa de células únicas e imagens (SCIMAP) do Instituto de Câncer Goodman da Universidade McGill, pelo processamento, marcação e aquisição de amostras para imagens de CMI; e a Brittney Armitage-Brown, do Serviço de Cuidados com Animais da Universidade Queen's, pela reprodução dos camundongos utilizados neste estudo. Este trabalho foi financiado pelos Institutos Canadenses de Pesquisa em Saúde (CIHR-394340) para CT e MK. Esta pesquisa foi financiada pelos Institutos Canadenses de Pesquisa em Saúde (CIHR).
Ética
Os experimentos descritos neste trabalho foram aprovados pelo Comitê Institucional de Cuidados com Animais da Queen's University (protocolo nº 2021-2228), de acordo com as diretrizes fornecidas pelo Conselho Canadense de Cuidados com Animais.
Você pode citar todas as versões usando o DOI https://doi.org/10.7554/eLife.96523 . Este DOI representa todas as versões e sempre será resolvido para a mais recente.
Este artigo é distribuído sob os termos da Licença de Atribuição Creative Commons , que permite uso irrestrito e redistribuição desde que o autor e a fonte originais sejam creditados


.jpg)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Nenhum comentário:
Postar um comentário